|
|
АРИТМИЯ СЕРДЦА |
Нормальная и аномальная электрическая активность сердечных клеток


Влияние патологических состояний на потенциал покоя и потенциал действия сердечных клеток
Нарушения ритма сердца и нарушения проведения могут возникать в результате изменений электрических свойств сердечных волокон при заболевании миокарда. Для выяснения особенностей таких изменений недавно был проведен ряд исследований. Для регистрации электрической активности клеток в препаратах миокарда, полученных у больных и экспериментальных животных, использовались стеклянные микроэлектроды. Результаты подобных исследований часто коррелируют с данными, полученными при изучении нормальных миокардиальных тканей, помещенных в измененную внеклеточную среду с целью моделирования условий, существующих при некоторых заболеваниях сердца.
Потенциал покоя
При многих заболеваниях сердца, вызывающих аритмию, отмечается тенденция к деполяризации мембраны сердечных клеток. К моменту написания данной главы мембранные потенциалы были зарегистрированы в предсердных клетках препаратов, полученных при ревматических и врожденных заболеваниях [51] и кардиомиопатии [52], а также в клетках миокарда желудочков и в волокнах Пуркинье, изолированных из пораженных ишемией и инфарктом областей сердца [53—56]. Во всех случаях мембранный потенциал покоя оказался менее отрицательным, чем в клетках аналогичных участков здорового сердца (рис. 3.10).
Причины уменьшения потенциала покоя в каждом из этих случаев не до конца понятны, хотя ясно, что определенный вклад могут вносить несколько факторов. При их рассмотрении лучше всего использовать обозначения из уравнения Гольдмана, Ходжкина и Катца, которое, как было показано ранее, позволяет достаточно точно определить потенциал покоя в волокнах Пуркинье при самых разных внеклеточных концентрациях ионов калия:
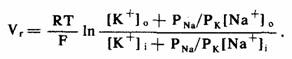
Если как типичные величины концентрации ионов и отношения проницаемости в этом уравнении принять [К+]0=4 мМ, [K+]i=150 мМ, [Na+]0=150 мМ, [Na+]i= 10 мМ и pna/pk=1/100, то можно показать, что в отсутствие значительных изменений температуры имеется 4 различных пути, позволяющих сделать потенциал покоя (Vr) менее отрицательным. Они таковы: 1) повысить внеклеточную концентрацию ионов калия — [К"+]0; 2) снизить внутриклеточную концентрацию ионов калия— [K+]i:: отношение pna/pk можно увеличить либо (3) путем повышения проницаемости мембраны для ионов натрия — pna, либо (4) путем снижения проницаемости для ионов калия — РK. Любое из этих изменений способно вызвать уменьшение потенциала покоя, но в поврежденных клетках может одновременно наблюдаться несколько таких изменений. Например, любое патологическое состояние, приводящее к ослаблению активности натриевого насоса, следует рассматривать как фактор, способствующий деполяризации поврежденных клеток, обусловленной, вероятно, как повышением [К+]0, так и снижением [К+]r. Нормальная потеря клеточного К+ и накопление Na+, сопровождающие электрическую активность (и наблюдающиеся в меньшей степени даже в покое), не являются, следовательно, легкоустранимыми (как это обычно бывает) посредством натриевого насоса. Другими словами, имеет место непрерывная суммарная потеря К+ из клеток (клеточный К+ эффективно замещается Na+), поэтому [К+]i постепенно уменьшается. Поскольку диффузия ионов из внеклеточного пространства в определенной степени ограничена и, следовательно, замедлена, в нем происходит накопление потерянных клетками ионов калия, что приводит к повышению [К+]0. Как отмечалось выше, и снижение [К+]i, и повышение [К+]0 могут способствовать изменению мембранного потенциала покоя.
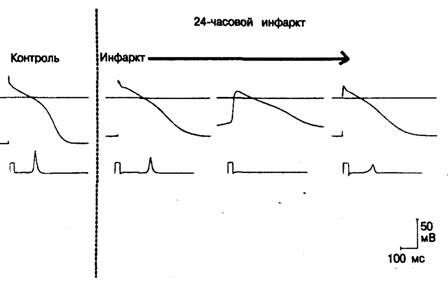
Рис. 3.10. Изменения потенциала действия, зарегистрированные в волокнах Пуркинье при инфаркте в сердце собаки.
В нижней части каждого фрагмента (записано на более высокой развертке) показано дифференцированное нарастание потенциала действия, которому предшествует дифференцированный калибровочный сигнал при скорости деполяризации 200 В/с; калибровочный импульс дифференциатора имеет форму прямоугольника. На левом фрагменте (контроль) — потенциал действия, зарегистрированный с волокна Пуркинье за пределами инфарктной зоны; отмечаются нормальный диастолический максимум (или потенциал покоя) и быстрое нарастание. Справа— потенциалы действия, зарегистрированные в трех различных волокнах Пуркинье на эндокардиальной поверхности в инфарктной зоне. Отметьте, что максимальный диастолический потенциал, амплитуда потенциала действия и скорость нарастания (Vmax) в зоне инфаркта ниже, чем в контрольной области. Угнетение потенциала покоя и потенциала действия особенно выражено на среднем фрагменте [53}.
Аналогично тому, как изменения [К+]0 и [К+]i могут одновременно способствовать снижению потенциала покоя, изменения PNa и РK, вносящие дополнительный вклад в деполяризацию мембраны, вероятнее всего, происходят одновременно. Например, предположим, что определенное патологическое состояние связано с увеличением потерь ионов Na через клеточную мембрану (иначе говоря, pna повышено). В этом случае отношение РNа— рк увеличится и (как следует из приведенного выше уравнения) потенциал покоя уменьшится. Поскольку такая деполяризация может происходить без каких-либо значительных изменений [К+]0 или [К+]i, потенциал равновесия К+ не изменится. Однако направленная наружу движущая сила для К+, равная Vm—ЕK, будет теперь больше нормы и (как результат направленного внутрь выпрямления) проводимость К+, а значит, и коэффициент проницаемости для К+ (РK) будут меньше нормы.
Постоянная деполяризация мембраны в этих условиях действительно связана с увеличением отношения pna—pk, однако такому изменению способствует не только повышение pna но и снижение РK.
С другой стороны, если какое-то заболевание сердца приводит к специфическому снижению РK (например, в результате изменений химического состава мембраны, обусловленных нарушением метаболизма белков или жиров), отношение pna—рк увеличится и возникнет деполяризация, несмотря на отсутствие изменений pna. Следует подчеркнуть, что такого специфического изменения рк пока не удалось выявить в пораженных сердечных клетках, хотя аналогичный эффект может быть искусственно вызван в изолированных миокардиальных тканях путем добавления в перфузионный раствор нескольких миллимолей цезия [57]. Несомненно, поддерживаемое увеличение потерь Na+ через клеточную мембрану в конце концов привело бы к изменению распределения катионов относительно поверхности мембраны, если бы натриевый насос не обладал способностью противостоять возрастающим потокам Na+ и К+. В ответ на относительное повышение проницаемости для натрия возникнет начальная деполяризация (как это описано выше), но затем будет постепенно развиваться повторная деполяризация, отражающая снижение внутриклеточной концентрации К+. Это связано с выходом К+ из клетки по мере поступления в нее Na+ для сохранения электронейтральности клеточного объема. Несмотря на то что проницаемость мембраны для ионов хлора в сердечных клетках обычно довольно низка, некоторое количество ионов хлора может поступать в клетки вместе с Na+ и водой, вследствие чего клетка слегка набухает, что соответственно приводит к дальнейшему уменьшению [К+]i.. Поскольку активность натриевого насоса определяется преимущественно уровнем [Na+]i с повышением последнего в таких клетках с утечкой она возрастает. Однако если пассивные потоки Na и К велики или активность насоса каким-либо образом ослаблена, то, несмотря на активацию работы насоса внутриклеточными ионами натрия, отмечаются значительные изменения уровня [Na+]i и [К+]i.
Нулевая фаза деполяризации
Для большинства изученных патологических состояний характерно замедленное нарастание с пониженной амплитудой потенциала действия (фаза 0) в волокнах Пуркинье или в клетках рабочего миокарда предсердий или желудочков [51—56, 58]. Такие изменения, по-видимому, во многом обусловлены" уменьшением мембранного потенциала пораженных клеток, хотя аналогичные изменения могут быть также следствием специфических для данного заболевания отклонений в предшествующих механизмах проводимости при отсутствии каких-либо изменений потенциала покоя. Мы пока не располагаем достаточной информацией относительно первичного специфического влияния заболеваний на механизмы проводимости перед возникновением направленного внутрь потока, вызывающего нарастание потенциала действия.
Ранее уже описывались механизмы инактивации быстрой натриевой проводимости при продолжительной деполяризации мембраны (после нарастания потенциала действия), а также отмечалась необходимость снижения реполяризации мембраны до уровня глубокого отрицательного потенциала (т. е. реполяризация потенциала действия) для полного устранения этой инактивации. Если мембрана реполяризуется не полностью, то в данных условиях инактивация устраняется лишь частично. Таким образом, после полной активации натриевой проводимости вполне достаточно гиперполяризации до уровня примерно —100 мВ, чтобы полностью устранить инактивацию и вернуть всем натриевым каналам способность к реактивации при следующем деполяризующем стимуле; при длительной же реполяризации до уровня от —60 до —70 мВ около 50 % натриевых каналов инактивируются и, следовательно, становятся недоступными для реактивации во время следующей деполяризующей стимуляции. При потенциале около —50 мВ практически все натриевые каналы остаются инактивированными и недоступными для немедленной реактивации (см. рис. 3.3).
Таким образом, в сердечных клетках, деполяризованных вследствие заболевания, только часть быстрых натриевых каналов доступна для входящего тока. В этом случае величина суммарного входящего тока во время фазы 0 потенциала действия уменьшается, а, значит, скорость нарастания и амплитуда снижаются (см. рис. 3.10). Такие потенциалы действия, нарастание которых зависит от входящего тока, текущего через частично инактивированные натриевые каналы, иногда называют «угнетенными быстрыми ответами» [16] в отличие от «медленных ответов», которые также имеют медленное нарастание, но зависят от входящего тока, протекающего по совершенно другим, фармакологи чески отличным мембранным каналам (см. выше). Из-за медленного нарастания и невысокой амплитуды скорость распространения угнетенных быстрых ответов существенно снижена. Например, скорость проведения потенциалов действия волокон Пуркинье может снизиться с 2—4 м/с до менее чем 0,5 м/с вследствие устойчивого состояния инактивации натриевых каналов, обусловленного деполяризацией мембраны. Дальнейшая деполяризация и инактивация натриевых каналов может привести к полной невозбудимости клеток, так что здесь возможно развитие блока проведения. Но хотя проводимость быстрых натриевых каналов может быть полностью инактивирована на уровне примерно —50 мВ, каналы медленного входящего тока (Na+/Ca2+-кaнaлы) становятся доступными для активации при потенциале ниже этого уровня [20, 21]. Следовательно, в этих условиях сильный деполяризующий стимул способен вызвать медленный входящий ток. Возможность этого обычно небольшого медленного входящего тока вызывать регенеративную деполяризацию, характерную для медленно распространяющегося потенциала действия, зависит от относительной величины мембранной проводимости К+. Как уже отмечалось при обсуждении потенциала покоя, деполяризация мембраны, вызванная, например, некоторым повышением ее проницаемости для Na+, способствует снижению проводимости для К+, что связано с наличием направленного внутрь выпрямления. В этих условиях медленный входящий ток может оказаться достаточным для инициации медленного ответного потенциала действия (см. рис. 3.3). С другой стороны, деполяризация мембраны, являющаяся следствием повышения [К+]0, связана с увеличением калиевой проводимости, так что в этом случае такой же небольшой медленный входящий ток может вызвать лишь незначительную деполяризацию. Однако, если медленный входящий ток возрастает, например, в присутствии катехоламинов, медленный потенциал действия может быть вызван и в условиях повышенной [К+]0 [59]. Из-за медленного нарастания скорость проведения медленных потенциалов действия очень низка. Таким образом, в результате сильной деполяризации мембраны скорость распространения потенциалов действия в волокнах Пуркинье может снизиться до величины менее 0,1 м/с [20].
Если деполяризация в фазу 0 замедляется до критического уровня, может возникнуть однонаправленный блок проведения [1]. В пучках предсердных или желудочковых волокон, а также в волокнах Пуркинье, стимулируемых с того или другого конца для инициации нормального потенциала действия, импульс проводится примерно с равной скоростью в обоих направлениях вдоль пучка. При критическом уровне подавления скорости нарастания потенциала действия проведение в одном направлении может исчезнуть, но будет медленно осуществляться в другом направлении (рис. 3.11). Критический уровень угнетения в различных областях сердца варьирует и частично зависит от геометрии сердечного синцития. Дальнейшее подавление нарастания потенциала действия и уменьшение амплитуды обычно приводит к блокированию проведения в обоих направлениях. Медленное проведение или однонаправленный блок проведения может возникнуть в пучках волокон, в которых отмечается либо угнетенный быстрый ответ, либо медленный потенциал действия.
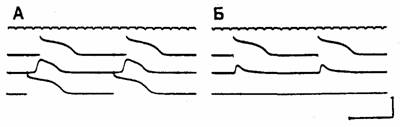
Рис. 3.11. Однонаправленный блок проведения в пучке волокон Пуркинье у собаки.
На верхней линии отмечено время с интервалами в 100 мс. Три расположенные ниже кривые — регистрация потенциала действия в трех различных клетках вдоль пучка волокон Пуркинье. Потенциалы действия на верхней кривой записаны с ближнего конца пучка, на средней — с центральной части пучка и на нижней — с дальнего конца пучка. Клетки центрального сегмента пучка были деполяризованы путем перфузии раствором с высоким содержанием [К'^] а поэтому потенциалы действия здесь имеют низкую амплитуду и замедленное нарастание. Фрагмент А: запись при стимуляции дальнего конца пучка. Импульс регистрируется сначала в клетке дальнего конца пучка (нижняя кривая), затем он проходит через центральный сегмент (кривая в середине) и, наконец, возбуждает клетки ближнего пучка (верхняя кривая). Фрагмент Б: запись при стимуляции ближнего конца пучка. Клетка, активность которой зарегистрирована на верхней кривой, возбуждается, поскольку она находится вблизи места стимуляции, однако проведение блокируется в угнетенном центральном сегменте (кривая в середине), поэтому активации дальнего конца волокна (нижняя кривая) не происходит [1].
Так как снижение мембранного потенциала в пораженных областях сердца вряд ли происходит совершенно одинаково, выраженность инактивации натриевых каналов скорее всего будет различной — от незначительной (импульсы проводятся быстро в виде быстрых потенциалов действия) и умеренной (импульсы проводятся относительно медленно как угнетенные быстрые ответы) до полной (импульсы, если они возникают, распространяются очень медленно как медленные потенциалы действия).
|
|
|
К содержанию книги: АРИТМИИ СЕРДЦА. Механизмы, диагностика, лечение
|
Последние добавления:
Виноградский. МИКРОБИОЛОГИЯ ПОЧВЫ
Ферсман. Химия Земли и Космоса
Перельман. Биокосные системы Земли
Вильямс. Травопольная система земледелия