|
|
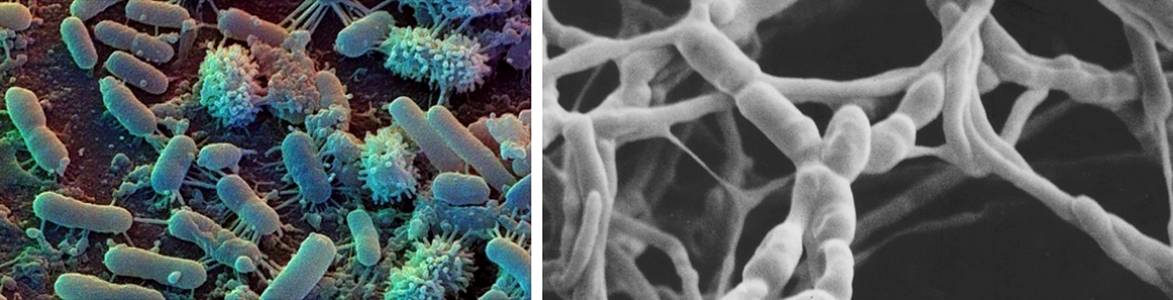
Почвенные организмы |
Зоомикробиотические комплексы в биогеоценозах
|
Смотрите также:
Мейен - Из истории растительных династий
Биографии биологов, почвоведов
|
И. В. СТЕБАЕВ Новосибирский государственный университет, Биологический институт
Почвенно-биологический подход к анализу биогеоценозов
Исследование биомасс имеет подчиненное значение. Более важно установить прямое и косвенное влияние животных и микроорганизмов на первичную продукцию, в том числе и идущее опосредованно через почву,— так уже пятнадцать лет тому назад М. С. Ги- ляров (1968) определил характерную особенность почвенно-биологического подхода к анализу биогеоценозов. Это особенно важно для первичных коисументов.
Действительно, для круговорота веществ, обеспечивающего восстановление первичной продукции, масса тел травоядных животных менее важна, чем, например, масса производимых ими экскрементов. Это особенно существенно для оценки роли беспозвоночных фито- и сапрофагов, так как у них коэффициент утилизации пищи (Ку = 27—37%) в 2—2,5 раза меньше, чем у растительноядных позвоночных. В результате масса экскрементов, продуцируемых за лето одними только саранчовыми в южносибирских степях, оказывается в четыре раза больше вторичной продукции тел самих насекомых. Абсолютное же значение этой массы так велико, что она составляет около трети ежегодного прихода обычного надземного растительного опада (Стебаев, 1968; Stebaev et al., 1968). Но самое важное состоит в том, что экскременты, образующиеся при малой степени утилизации, представляют прекрасную базу для бурной и многообразной деятельности микроорганизмов, на что впервые еще в 1887 г. обратил внимание И. И. Мечников при своих исследованиях в украинских степях.
Объективные данные по этому вопросу появились около 20 лет назад в Сибири в начале по отношению к почвенным сапрофагам (Стебаев, 1958; Козловская, 1964, 1967), а затем и к сапрофагам, обитающим на поверхности почвы, и к фитофагам, обитающим над почвой в травостое (Стебаев, Гукасян, 1963; Стебаев и др., 1964; Stebaev et al., 1968). Вскоре аналогичные сведения стали известны и для обитателей древесных крон (Злотин, Ходашова, 1974). Выяснилось, что уже в передних отделах кишечных трактов насекомых происходит бурное развитие в первую очередь обычной почвенной микрофлоры, сопровождаемой, конечно, также и специфической кишечной. У сапрофагов своеобразие состояло в сокращении количества грибов и в увеличении бактерий. В первую очередь это касалось бацилл, обычно учитываемых на МПА и КАА. Дальнейшее развитие этой тенденции было прослежено и в экскрементах. Все это говорит о том, что при переработке пищи насекомыми происходит форсирование микробиотической сукцессии, обычной для почв и подстилок. В случае сапрофагии эта сукцессия начинается, по-видимому, еще в субстрате, служащем им пищей.
Фитофаги же, обитающие в травостое, показали себя в этих исследованиях как возбудители особой микробной сукцессии, начало которой кладется стремительным развитием целлюлозоразрушающих бактерий.
Очевидно, и в том и в другом случае речь должна идти о ферментативном синергизме обычных почвенных микроорганизмов, поглощенных животными с пищей, и самих этих животных, что и было показано в недавнее время на биохимическом уровне Б. Р. Стригано- вой (1980). В результате сложилось представление о так называемом зоомикробиотическом комплексе (ЗМК) как об особом функциональном блоке биогеоценоза, имеющем в общем симбиотическую природу (Стебаев, 1968). Не исключено, на что не раз обращал внимание в своих лекциях Н. А. Красильников, что регулярный пассаж части популяций свободноживущих микроорганизмов через тела и экскременты животных является даже непременным условием поддержания популяций микроорганизмов в ландшафте. Другими словами, у них может быть установлена, пусть даже факультативная, зоофилия, подобная таковой у цветковых растений.
Функционирование ЗМК определяется, на наш взгляд, по крайней мере шестью звеньями, которые превращают часть биотического круговорота веществ, идущую через ЗМК, в наиболее форсированную и управляемую регуляторными механизмами. Рассмотрим эти звенья в основном на примере лучше изученных в этом отношении степных саранчовых и чернотелок ( 1, 1—6).
Звено управления круговоротом путем отыскивания наиболее эффективного для минерализации объекта в фитомассе ( 1, 1). Это звено выявляется в трофоэтологических исследованиях, которые могут установить: полноту трофического контроля насекомых над всей площадью растительной ассоциации; вероятность отыскания групп форм, являющихся мацераторами этих тканей и разрушителей . люлозных оболочек ( 3). В результате происходит бурное высвобождение внутриклеточного азота и быстрая его минерализация.
Таким образом, в подчас еще зеленой растительной пищевой массе в кишечнике насекомых, находящихся в травостое, возбуждается и быстро реализуется микробиотическая сукцессия, обычно протекающая в подстилке ш в почве и притом весьма длительно. Есть ост нование полагать, что решающую роль здесь играет синэргическое действие целлюлозоразрушаннцих ферментов как бактериального, так и животного происхождения (Ненюков, Парфентьев, 1929; Стри- ганова, 1970, 1980). Последнее имеет особое значение, так как именно медленное развитие целлюлозоразрушающих фаз в детритных субстратах задерживает начало разрушения растительного опада минимум на два—четыре дня (Кононова, 1963), а в природной степной подстилке — па два-три года (Семеиова-Тян-Шанская, 1960). В то же время глубокая и быстрая минерализация, конечно, уменьшает шансы образования гумусовых кислот, однако не нужно забывать, что фенольные метаболиты разрушенной клетчатки создают структурные элементы молекул именно этих кислот.
Обитатели подстилки сапрофаги жуки-чернотелки (Стебаев и др. 1964; Stebaev et al., 1968) также приближают микробиотическую сукцессию к финальным стадиям, но уже к тем, которые обычны не и подстилке, а в самой почве ( 3). Они обеспечивают эти стадии в большей степени, чем саранчовые, так как их экскременты отличаются обычно в 0,5—2 раза большим КМ и в них постоянно присутствуют актиномицеты. Общий характер микрофлоры в ^свежих экскрементах чернотелок ближе к таковому в почве, чем в подстилке. В то же время их нельзя назвать начальными возбудителями почвенной сукцессии — они скорее лишь ускорители и продолжатели той ее стадии, которая самостоятельно начинается в растительных остатках подстилки еще до их поедания чернотелками. Об этом говорит близкое к таковому в подстилке общее число микроорганизмов, а также в целом небольшое количество целлюлозоразрушающих и почти полное отсутствие флюоресцентных форм, как известно, типичных для самых ранних стадий микробных сукцессий.
Как признак в основном лишь ускоренного окончания обычной микробиотической сукцессии следует рассматривать и общий характер как кишечной, так и экскрементной микрофлоры у постоянных обитателей самой почвы (Козловская, Жданникова, 1961, 1962; Козловская, 1964, 1967). Правда, у некоторых из них как признак зоо- гепного оживления микрофлоры обнаруживается большее, чем в подстилке обилие целлюлозоразрушающих форм, что, возможно, связано с поеданием живых корней. В то же время показанное JI. С. Козловской (1967) и JI. С. Козловской совместно с И. Н. Раковой (1978) обилие в экскрементах таких беспозвоночных мобилизованных форм органогенов ясно говорит о важности трофической деятельности этих обитателей ризосфер для улучшения условий роста растений.
Таким образом, характер изменения микробной активности в почвенной части зоомикробиотического комплекса подобен таковому в профиле почвы (Мишустин, 1954, 1975).
3. Звено управления круговоротом путем коррекции развития микрофлоры в зоомикробиотическом комплексе со стороны популя- ционных и почвенных факторов ( 1, 4). Половая структура популяций насекомых может влиять на активность ЗМК. Суммарный вес самок саранчовых и массы производимых ими экскрементов во всех популяциях в полтора—два раза больше, чем у самцов. Однако это еще не означает, что именно самки играют ведущую роль в ЗМК, так как уровень метаболизма у них в 1,5—2 раза ниже, чем у самцов.
В тех случаях, когда морфологический поведенческий половой диморфизм выражен хорошо, самцы как более активные особи оказываются носителями более обильной микрофлоры ( 4). Будучи значительно более подвижными, чем самки, они также играют большую роль и в распространении микрофлоры с экскрементами по всей площади биогеоценоза.
Важна и степень, так сказать, экологического процветания популяции. На примере трех видов саранчовых видно, что общее количество микроорганизмов в их экскрементах тем выше, чем больше численность насекомых. Аналогичное явление описано Л. С. Козловской (1967) и для дождевых червей. Действие такого популяци- онного фактора, как это видно на рисунке, не легко отделить от влияния микробиотического фона почв, из которых черпается микрофлора кишечных трактов и куда она в конце концов возвращается с экскрементами.
Анализ различий в составе микрофлоры, почв и экскрементов все же показывает, что фоновые различия проявляются в экскрементах одного вида на разных почвах, главным образом за счет изменения обилия и состава олигонитрофилов. В целом здесь в соответствии с известной концепцией М. С. Гилярова (1954) мы находим пример того, что в биогеоценозе действует не просто вид вообще, а качественно специфичная именно для данного биогеоценоза часть вида — популяция. Причем обычно трудно выявляющаяся непосредственным путем физиологическая специфика популяций в данном случае хорошо проявляется в функциональных особенностях микрофлоры ЗМК, сопровождающей популяцию.
4. Звено управления круговоротом путем повышения эффективности усвоения растениями конечных продуктов деятельности ЗМК ( 1, 5) осуществляется путем микробогенного образования в экскрементах биологически активных веществ типа биотина, стимулирующих рост и прорастание растений, на наличие которых у насекомых давно обратил внимание М. С. Гиляров (1958). Повышенное содержание подобных веществ в экскрементах педобионтов в лабораторных условиях показано JL С. Козловской (1967) и С. Р. Диком (1967), а для саранчовых в поле — нами (Stebaev et al., 1968). С помощью биотеста (Saccharomices cerevisiclae) было выяснено, что за два—четыре часа пребывания растительной массы в кишечном тракте саранчовых содержание биотина у разных видов возрастает в три—двенадцать раз, причем у видов, питающихся дерновинными злаками, это проявляется особенно сильно.
5. Звено управления круговоротом с помощью коррекции места поступления экскрементов в почву { 1, 6). В экскрементах саранчовых, попавших в почву, через 5 дней обилие целлюлозоразру- шающих и других характерных для них форм бактерий сильно сокращается (Stebaev et al., 1968). Спектр микробного населения экскрементов, давно лежащих на поверхности почвы, уже мало отличается от такового в экскрементах подстилочных сапрофагов-чернотелок и приближается к таковому в местной почве ( 3). Не исключено, что убывает и количество накопленных в них органогенов. В то же время известно, что свежие экскременты, попадая в подстилку, могут резко повышать ее общую микробиотическую активность (Зло- тин, Ходашова, 1974). Тем более важно местонахождение экскрементов на почве именно с первых минут их поступления.
Хронометрирование движения саранчовых показало, что более 60% времени они проводят в покое внутри дерновинных злаков и кустиков полыни, где, по-видимому, :в основном и происходит дефекация. Действительно, микрокартирование положения экскрементов на поверхности почвы показало, что они концентрируются именно внутри дерновин ( 6). Чем разреженнее травостой, т. е. чем меньше вероятность попадания экскрементов в область ризосфер, тем оказывается лучше выражено их дискретное распределение и узловая концентрация у корневых шеек растений. Это соответствует хронометрическим данным, показывающим, что в такой ситуации связь самих насекомых с растениями как с микроубежищами проявляется особенно сильно.
Сходным образом массы экскрементов чернотелок концентрируются под кустами карагана (Мордкович, 1973). Аналогичное явление обнаружено и для экскрементов крупных педобионтов (Козловская, 1967). Последним автором показано, что вертикальные миграции беспозвоночных в почве обеспечивают инокуляцию активной микрофлоры в более глубокие горизонты. Известно также, что в почве, из которой удалены личинки насекомых, микробная активность сильно снижается (Witkamp, Groslei, 1966).
Таким образом, гранулы экскрементов, богатые микрофлорой, органогенами и биостимуляторами растений, как бы направляются самими насекомыми к корневым шейкам растений. Это в свою очередь должно повышать эффективность восстановления биомассы этих растений, отчужденной насекомыми.
6. Звено управления круговоротом с помощью изменения сроков поступления в почву экскрементов как зоомикробиотически измененной части растительного опада особо проявляется в деятельности надпочвенной части ЗМК. Обычный листовый опад обеднен питательными веществами, на почве он оказывается лишь осенью, когда вследствие недостатка тепла, а в условиях континентального климата и влаги, он разлагается медленно и соответственно медленно и мало способствует пополнению запаса органогенов в почве. Экскременты листогрызущих насекомых можно рассматривать как некий зооген- ный листовый опад, состоящий из растительных тканей, богатых органогенами и начинающий разлагаться еще на своем пути к земле. Срок его образования определяется фенологией насекомых, а ле растений. Важно, что он образуется не осенью, а в середине лета, когда нет недостатка в тепле, а в континентальных районах — и во влаге (Носин, 1963). Вследствие этого такой «опад» может быть вполне сопоставим с обычным по своему значению для круговорота, несмотря на то, что, по-видимому, аналогичный эффект проявляется и в других ярусных частях ЗМК, особенно в подстилочной.
Значение зоомикробиотического комплекса в разных зональных типах биогеоценозов
Значение зоомикробиотического комплекса в разных зональных типах биогеоценозов не одинаково по ряду причин, среди которых имеет значение и смена видового состава насекомых. По-видимому, везде наиболее активными стимуляторами микрофлоры являются виды, связанные с растениями-эдификаторами: в степях — с дерно- винными злаками, в полупустынях — с полынями (см. таблицу). Учет микрофлоры экскрементов разных насекомых и других показателей (Стебаев, 1968) позволяет проследить самые общие закономерности ( 7).
Зоомикробиотический комплекс биогеоценозов ЗМК
В зональном ряду биогеоценозов почвенная часть ЗМК играет наибольшую роль в черноземных степях, быстро убывая при переходе к каштановым почвам. В биогеоценозах с этими почвами ведущую роль играет напочвенная часть ЗМК (чернотелки) и надпочвенная (саранчовые). В биогеоценозах со светлокаштановыми почвами роль надпочвенной ЗМК возрастает особенно сильно. Это означает, что в сухостепных и пустынных биогеоценозах ход почвенных микробиотических процессов, обеспечивающих восстановление Первичной продукции, во многом решается в зеленом ярусе биогеоценоза. Почвенная же часть ЗМК, несмотря на тяготение многих насекомых к почве (Гиляров, 1951), может сохранять здесь лишь подчиненную роль (Стриганова, 1980).
Итак зоомикробиотический комплекс биогеоценозов (ЗМК) — продукт длительной коэволюции растений, животных и микроорганизмов. Он обеспечивает кооперирование этих организмов в использовании ресурса, а также ускоренное возобновление круговорота органогенов и вертикальной миграции веществ в биогеоценозах.
В основе его эволюции должна была лежать первичная бактериофагия беспозвоночных. На ее основе могла возникнуть облигат- ная для животных синергическая связь с микроорганизмами. Суть механизмов поддержания ЗМК — в эффекте сочетания сильных ферментативных систем микроорганизмов и совершенной пищевой ориентации поведения насекомых, а также в отзывчивости растений на мобилизированные животными и микроорганизмами формы органогенов и на образованные ими же биологически активные вещества. Наибольшее значение этот комплекс должен иметь в сухостепных ландшафтах с континентальным гидрохимическим режимом. Описанные явления показывают необходимость осторожности1 при отнесений насекомых-фитофагов к безусловно вредным для сельского хозяйства, учитывая важность охраны животного населения почв, подстилки и трав как носителей полезной микрофлоры.
Целесообразной была бы выработка мер культивирования зоомикробиотических комплексов на агроугодьях, в том числе и такой части комплексов, как саранчовые, которые играют в природе не меньшую, если, не большую, положительную роль, чем дикие копытные и опылители, при этом не исчезая, в отличие от этих животных, даже на сильно измененных сельскохозяйственных землях.
ЛИТЕРАТУРА Биология океана / Под ред. М. С. Виноградова. М.: Наука, 1977. Т. 2. 398 с. Гиляров М. С. Использование насекомыми почвенного яруса в сухих частях ареалов.— Успехи соврем, биологии, 1951, т. 31, вып. 2, с. 161—169. Гиляров М. С. Вид, популяция, биогеоценоз.— Зоол. журн., 1954, т. 33, вып. 4, с. 769—778. Гиляров М. С. Биологически активные вещества, выделяемые насекомыми.— Успехи соврем, биологии, 1958, т. 11, вып. 2(5), с. 208—216. Гиляров М. С. Почвенный ярус биоценозов суши.— Успехи соврем, биологии, 1968, т. 66, вып. 1(4), с. 121—135. Лип С. Р. К вопросу о содержании витаминов «В» в почвах и экскрементах беспозвоночных.— В кн.: Взаимодействие леса и болота. М.: Наука, 1967, с. 98— 104. Злотин Р. ИХодашова К. С. Роль животных в биологическом круговороте лесостепных экосистем. М.: Наука, 1974. 82 с. Козлов А. Т., Свешников В. А. Поведение личинок ручейников Neureclipsis bi- maculata в условиях эксперимента.— В кн.: Управление поведением животных. М.: Наука, 1977. 147 с. Козлов А. Т.у Черемисина В. Т. Строительное поведение личинок ручейников Phryganea striata в условиях эксперимента.— В кн.: Материалы второго Все- союз. симпоз. по поведению водных беспозвоночных. Борок, 1975, с. 42—44. Козловская Л. С. Роль почвенных организмов в разложении растительных остатков в торфе.— В кн.: Докл. сибирских почвоведов к VIII Междунар. почв, конгр. Новосибирск: Наука, 1964, с. 78—84. Козловская Л. С. Роль почвенных беспозвоночных в трансформации органического вещества болотных почв. Л.: Наука, 1967. 243 с. Козловская Л. С., Жданникова Е. Н. Совместная деятельность дождевых червей и микрофлоры в лесных почвах.— Докл. АН СССР, 1961, т. 139, № 2, с. 470— 473. Козловская Л. СЖданникова Е. Н. Взаимодействие почвенной фауны и микрофлоры.—Изв. СО АН СССР, 1962, № 5, с. 107—117. Козловская Л. С., Ракова И. Н. Ферментативная активность экскрементов беспозвоночных.— В кн.: Исследования по лесному болотоведению и мелиорации. Петрозаводск, 1978, с. 64—80. Кононова И. М. Органическое вещество почвы. М.: Изд-во АН СССР, 1963. 314 с. Мечников И. И. Замечание о хлебном жуке.— Сел. хоз-во и лесоводство, 1887, № 134, с. 231—249. Мишустин Е. Н. Закон зональности и учение о микробных ассоциациях почвы.— Успехи соврем, биологии, 1954, т. 37, вып. 1, с. 1—21. Мишустин Е. Н. Ассоциации почвенных микроорганизмов. М.: Наука, 1975. 164 с. Мордкович В. Г. Устойчивость и динамика агрегаций педобионтов под крупными растениями Курайской степи.— В кн.: Экология и структура населения почвообитающих животных Алтая: Вопросы экологии. Новосибирск, 1973, с. 182—194. Ненюков Д. В.} Парфентьев А. И. Пищеварительные процессы в связи со строением кишечника у перелетной саранчи.— Защита растений, 1929, № 6, с. 21—37. Носин В. А. Почвы Тувы. М.: Изд-во АН СССР, 1963. 342 с. Плеханов Г. Ф., Купресова. В. БТрофимов Л. Г., Романенко В. Н. Поведение клеща Hyaloma asiaticus P. Sch. et E. Schl. при поиске объектов питания.— В кн.: Этология насекомых и клещей. Томск: Изд-во Том. ун-та, 1977, с. 87— 108. Семенова-Тян-Шанская А. М. Динамика накопления и разложения мертвых растительных остатков в лугово-степных и луговых ценозах.— Ботан. журн., 1960, т. 45, с. 1342—1350. Стебаев И. В. Роль почвенных беспозвоночных в развитии микрофлоры почв Субарктики (на примере личинок Tipulidae, Diptera).— Докл. АН СССР, 1958, т. 122, № 4, с. 720—722. Стебаев И. В. Характеристика надпочвенного и почвенного зоомикробиотиче- ских комплексов степных ландшафтов Западной и Средней Сибири.— Зоол. журн., 1968, т. 47, вып. 5, с. 661—675. Стебаев И. В., Гукасян А. Б. Прямокрылые насекомые Tettigonioidae, Acrididae как стимуляторы микробиологических процессов распада и минерализации растительной массы в луговых степях Западной Сибири.— Зоол. журн., 1963, т. 42, вып. 2, с. 216—221. Стебаев И. В., Наплекова Н. ИГукасян А. Б. Саранчовые (Acrididae) и чернотелки (Tenebrionidae) как стимуляторы микробиологических процессов в почвах сухих степей Тувинской автономной республики.— Почвоведение, 1964, № 9, с. 89-95. Стебаев И. В., Пшеницина. Л. Б. Избирательность питания доминантных видов саранчовых прииртышских степей и пойменных лугов, определяемая методом диагностики ботанического состава экскрементов.— В кн.: Вопросы экологии (вид, популяция, сообщества). Новосибирск, 1978, с. 18—58. Стебаев И. В., Соболев Н. Н., Лопаткин А. В. Анализ стациальной верности и расселения популяциониых групп саранчовых по микроландшафтам с помощью полевого эксперимента (в связи с вопросом о микроландшафтной ориентации насекомых и о нространственно-временной структуре их популяций, на примере Dociostaurus brevicollis Ev. Chorthippus albomarginatus De G.).— В кн.: Вопросы экологии. Новосибирск, 1978, с. 60—69. Стриганова Б. Р. О разложении целлюлозы в кишечнике кивсяков Pachyiulus foetidissimus (Mur.) (Julidae, Diplopoda).—Докл. АН СССР, 1970, т. 190, № 3, с. 703—705. Стриганова Б. Р. Питание почвенных сапрофагов. М.: Наука, 1980. 244 с. Шовен Р. Физиология насекомых. М.: Изд-во иностр. лит., 1980. 368 с. Stebaev J. V., Naplekova N. J., Wolkowincer V. V Epigaeische Zoomikrobio- ten-komplexe mit Orthopteren und Tenebrioniden im siidostlichen Altai — Ge- birge und ihre Beziehungen zu bodenbildenden Processen.— Pedobiologia, 1968, Bd. 8, S. 345—386. IVitkamp M., Grosley D. A. The role of arthropods and mikroflora in breakdown of White Oak Litter.— Pedobiologia, 1966, Bd. 6, S. 293—303. |











К содержанию книги: Почвенные организмы как компоненты биогеоценоза