|
|
Почвенные организмы |
Активность ферментов азотного обмена в почвах естественных фитоценозов
|
Смотрите также:
Мейен - Из истории растительных династий
Биографии биологов, почвоведов
|
Московский государственный университет им. М. В. Ломоносова
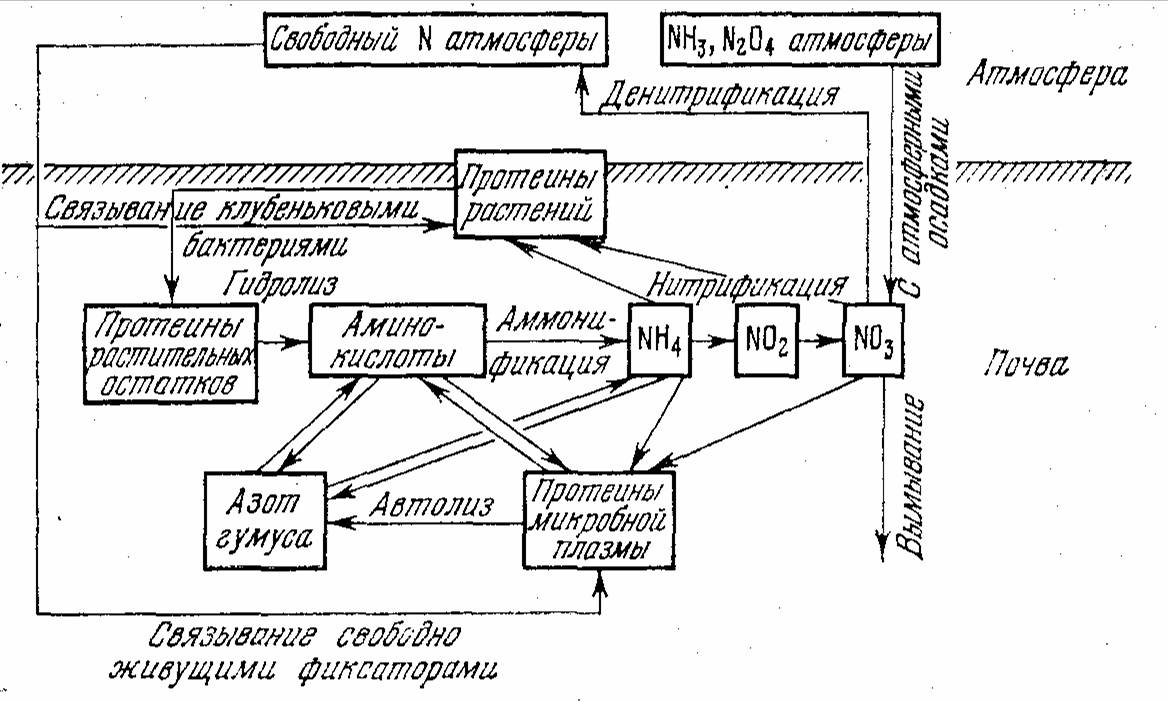
Азотный обмен в почвах
Наиболее дефицитным элементом в дерново-подзолистых почвах является азот. Поэтому большой научный и практический интерес представляет изучение путей его мобилизации. В трансформации азотсодержащих органических веществ, входящих в состав растительных и животных остатков, подстилок и почвы, существенная роль принадлежит почвенной микрофлоре, проявляющей свою функциональную активность через внутри- и внеклеточные ферменты, учет активности которых важен при изучении путей, оптимальных условий и расчетах круговорота азота, а также при поиске надежных способов диагностики азотного питания растений. Данные же по уровню ферментов азотного обмена в подзолистых и дерново-подзолистых почвах естественных фитоценозов чрезвычайно малочисленны (Раськова, 1978; Щербакова, 1979; Ивлева и др., 1981).
Пристальное внимание к изучению целинных почв связано с необходимостью познания закономерностей саморегулирующихся природных систем для правильного использования их в земледелии и контроля за изменениями, происходящими при этом в почвах. Поэтому вполне актуальным является изучение биологической активности целинных почв по интенсивности трансформации органических веществ, содержащих азот, по характеру профильного распределения ферментов азотного обмена, связи их с другими свойствами почв и растительностью.
Исследования проводили на опытных площадках в Малинском лесничестве Краснопахорского лесхоза Московской обл. Изучали дерново-подзолистые почвы, сформированные под липовым, березовым, еловым и сосновым лесами с преобладанием в травяном покрове осоки волосистой, подзолистую почву под ельником зеленомошным и дерновую под лугом.
В подстилках (А0) этих почв углерода содержалось от 26 до 34%, в гумусово-аккумулятивном горизонте (At) — от 2,4 до 5,3%, в переходном (AIA2) — от 1,3 до 1,6%; азота в А0 — от 1,07 до 1,65%; в At — от 0,19 до 0,32%, в AiA2 —от 0,06 до 0,12%. Многие химические, физические и биологические свойства почв этого региона описаны в литературе (Карпачевский, 1977; Раськова, Звягинцев, 1981).
При отборе образцов по профилю брали полный столбик почвы, не допуская разрыва между горизонтами, выделяемыми по морфологическим свойствам. Такой способ отбора позволяет получить полную характеристику генетических горизонтов и охарактеризовать суммарную величину активности в расчете на вес столбика почвы с определенной площадью поверхности.
Активность уреазы и протеазы
Активность протеазы определяли во влажных образцах, так как в воздушно-сухих она резко снижается. Подстилку освобождали от толстых сучьев, шишек, а затем размалывали на мельнице, получая однородную мелкоизмельчен- ную массу. Для анализа из подстилок брали навеску в 0,2 г, из гумусово-акку- мулятивного горизонта — 1 г, переходного — 2 г в расчете на абсолютно сухой образец. В качестве субстрата использовали казеин (10 мл 1%-ного раствора), растворение которого проводили при небольшом нагревании в 0,1-ном растворе NaOH; рН полученной смеси затем доводили до 7,4 фосфатным буфером.
Реакционную смесь инкубировали 21ч при 30° С при периодическом перемешивании, затем центрифугировали при 8 тыс. об/мин в течение 10 мин. К 5 мл супериатанта добавляли 2 мл 0,1 н. серной кислоты и 3 мл 20%-ного раствора сернокислого натрия. Осадившийся белок отфильтровывали. В алик- вотной части фильтрата определяли содержание аминокислот, используя нин- гидриновый реактив (1 г нингидрина растворяли в 37 мл этилового спирта, а затем добавляли 13 мл 4 н. ацетатного буфера; рН 5,5). Развитие окраски в реакционной смеси, содержащей фильтрат и нингидриновый реактив, протекало при 100° в течение 15 мин. После чего смесь разбавляли 50%-ным этанолом до определенного объема и измеряли оптическую плотность при 570 нм. Количество аминокислот определяли по стандартной кривой, построенной по глицину. Контролем служила реакционная смесь без субстрата; необходим контроль и на реактивы.
Активность уреазы определяли в воздушно-сухих образцах. Состав реакционной смеси готовили согласно прописи (А. Ш. Галстяна, 1974), количество аммиака определяли путем образования окрашенного комплекса с реактивом Несслера (Аринушкина, 1961); интенсивность накопления свободных аминокислот оценивали на аппликационных полотнах по методике, описанной ранее (Гельцер, Комовникова, 1979).
Показано, что уровень активности протеазы и уреазы в исследованном ряду почв существенно изменялся в течение вегетационного сезона (табл. 1). Наибольшую амплитуду сезонных колебаний наблюдали в подстилках, где активность протеазы и уреазы от одного периода наблюдений к другому изменялась в 2—4 раза. В гумусово-аккумулятивном и переходном горизонтах сезонные колебания могут быть значительными, но они несколько меньше, чем в подстилках. Видимо, биохимический состав подстилок, численность и активность микроорганизмов в них изменяется во времени сильнее, чем в минеральных горизонтах. Замечено, что нет строгой синхронности в сезонных изменениях активности ферментов в подстилках и минеральных горизонтах, а периоды максимальной их активности меняются в зависимости от гидротермических условий года. Активность протеазы в А0 под ельником волосистоосоковым в 1976 г., отличающимся пониженной температурой и повышенной влажностью, наибольшей была в июне (43,4 мг/г), а в 1977 г., с теплой и относительно засушливой погодой—1 октября (67,3 мг/г) (табл. 1).
Анализ индивидуальных образцов, отобранных в один срок, показал, что коэффициент пространственного варьирования (V) активности протеазы и уреазы в дерново-подзолистых почвах колеблется от 10 до 48%, примерно в таких же пределах он изменяется и в дерновой почве (табл. 2.) Неоднородность отмечена и для ряда физических и химических свойств этих почв (Карпачевский и др., 1968; Дмитриев, Самсонова, 1975).
Для дерново-подзолистых почв, сформированных под естественной растительностью, характерно резкое снижение содержания углерода и азота с глубиной. В подстилках, состоящих из растительных остатков различной степени разложения, содержится около 30% углерода и 1,15% азота, в гумусово-аккумулятивном горизонте в 8 раз меньше углерода (3,5%) и в 4,4 раза — азота (табл. 3). Аналогичным образом распределены по профилю ферменты, но активность протеазы изменяется более контрастно. Так, под действием протеазы, содержащейся в 1 г подстилки, образуется в среднем в 14 раз больше аминокислот, чем в горизонте At, и в 39 раз —чем в горизонте AiA2 (см. табл. 3). В бурых горно-лесных почвах под буко- во-пихтово-еловыми лесами отношение активности протеазы в подстилке к активности в верхнем минеральном горизонте значительно меньше (Чернявский, 1976).
Установлено, что в профиле дерново-подзолистых почв активность ферментов азотного обмена тесно коррелирует с содержанием гумуса и азота. Коэффициенты корреляции (г) активности ферментов с некоторыми химическими и физическими свойствами целинной дерново-подзолистой почвы достоверны на высоком уровне значимости (см. ниже). Активность ферментов сопряжена и с влажностью почв, причем для протеазы эта связь теснее, чем для уреазы.
Важно отметить также, что характер распределения активности ферментов по генетическим горизонтам аналогичен интенсивности накопления свободных аминокислот на аппликационных полотнах, содержание которых в верхней части профиля в 3—10 раз больше, чем на глубине 21—24 см (табл. 4). Корреляционная связь между активностью ферментов азотного обмена и содержанием углерода и азота обнаружена и для других почв (Хазиев, Агафарова, 1975; Ross, 1977; Колоскова, Мурзатова, 1978). Замечена связь протеоле- тической активности почв с интенсивностью биологического круговорота веществ, измеряемой величиной опадо-подстилочного коэффициента (Смольянинов и др., 1968).
Таким образом, несмотря на более высокую обогащенность подстилок углеродом, азотом и ферментами (в расчете на 1 г образца), содержание этих веществ в толще минеральных горизонтов с учетом их мощности (в расчете на вес горизонта с основанием в 1 см2) значительно выше, так на площадь в 1 см2 приходится в среднем лишь 0,055 г подстилки, а горизонты At и AtA2 на этой же площади весят по нескольку граммов.
Оценка генетических горизонтов и почв
Оценка генетических горизонтов и почв в целом по запасу ферментативных белков и биогенных элементов дает представление о характере их аккумуляции и о роли отдельных горизонтов в процессах почвенного метаболизма. Высокая ферментативная активность в подстилках и резкое снижение ее с глубиной отмечались многими исследователями, но при этом не определялось содержание ферментов (в единицах активности) во всей толще горизонта и их количественное соотношение. Однако несомненна значимость именно этих показателей при выявлении специфических особенностей почв разных биоценозов, установлении влияния факторов почвообразования на биологические свойства почв, а также при изучении оптимальных соотношений между ферментативным потенциалом и другими свойствами почв.
Важной характеристикой почв является степень обогащенностй их органического вещества ферментативными белками, о которой судят по количеству ферментативных единиц, приходящихся на грамм углерода. Данные табл. 3 и 5 убеждают в том, что больше обогащены протеазой подстилки под лесной и особенно под травянистой растительностью, чем минеральные горизонты. Это связано, видимо, с тем, что белки растительной массы опада стимулируют развитие определенных групп микроорганизмов, функциональная деятельность которых сопровождается усиленным синтезом протеаз. Уреазой органическое вещество всех горизонтов обогащено в одинаковой мере, за исключением нижних слоев подстилки сильноподзолистой почвы, где на грамм углерода приходится минимальное количество этого фермента (3—12 ед.).
Таким образом, показана пространственная и временная изменчивость уровня протеазы и уреазы, определены характер профильного распределения ферментов в почвах естественных биоценозов и соотношение их активности с содержанием гумуса, азота, интенсивностью накопления аминокислот и с влажностью почв.
Остановимся более подробно на специфических особенностях уровня активности ферментов, их динамики, характера распределения и суммарного содержания в почвах, сформированных под разными типами леса. Сезонная динамика активности протеазы и уреазы в верхних горизонтах иллюстрируется на примере трех биогеоценозов (см. табл. 1), почвы которых в наибольшей мере различаются между собой по активности ферментов в верхнем высокобиогенном слое (А0). Видимо, в подстилке дерновой почвы выше активность как протеазы, так и уреазы по сравнению с лесными фитоценозами, Обогащённость азотом и ферментами почв, сформированных под разными типами леса
Связано это, видимо, с тем, что опад под лугом и березовым лесом более богат зольными элементами и азотом (Карпачевский, 1977; Вайчис, Данусявичус, 1978), обильнее заселяется микрофлорой, что ведет и к более интенсивной его трансформации. В трехслойной подстилке сильноподзолистой почвы под ельником зеленомошным активность обоих ферментов резко снижается по мере увеличения степени ее минерализации и в подзолистом горизонте обнаруживаются лишь их следы (см. азотный обмен в почвах).
Таким образом, по величине активности протеазы и уреазы подстилки изученных фитоценозов располагаются в следующий ряд: луг > березняк ^ липняк > ельник ^ сосняк. В гумусово-аккумуля- тивном горизонте уровень уреазы выравнен во всех почвах. По активности протеазы в наибольшей мере различаются почвы под березовым и еловым лесами. Протеолитическая активность в почве под березовым лесом существенно меньше, чем под ельником (см. табл. 2 и 1). В течение сезона различия эти проявляются в разной степени.
Специфика почв разных фитоценозов выявляется и по соотношению содержания ферментов в генетических горизонтах при расчете активности на вес горизонта с площадью поверхности в 1 см2.
Из средних данных за 1976—77 гг. (табл. 5) видно, что в подстилках хвойных пород содержится 22% протеазы от суммарного ее запаса в 3 верхних горизонтах, а в подстилках лиственных пород — всего лишь 7 %. В сильноподзолистой почве основная часть протеазы и уреазы сосредоточена в подстилочных слоях, а в дерновой под лугом содержание их в подстилке составляет лишь 1—2%. Существенны различия и по содержанию ферментов в горизонте At. В почве под ельником волосистоосоковом гумусово-аккумулятивный горизонт почти в 3 раза меньше обогащен протеазой и в 4 раза уреазой, чем под березняком.
Величина активности на вес горизонта объединяет в себе информацию по активности фермента на 1 г образца и по величине соответствующего слоя, поэтому сравнение ферментативного потенциала почв, различающихся между собой по морфологическому строению профиля или по мощности одноименных горизонтов, необходимо проводить не только по активности на грамм почвы, но и по обогащенно- сти генетических горизонтов ферментами и расчете на вес горизонта.
Таким образом, приведенные данные достаточно убедительно показывают влияние растительности на формирование уровня и характера распределения ферментативного фонда почв по горизонтам (см. табл. 5; 1, а, б). Как ранее было показано для каталазы (Раськова и др., 1977), наиболее общей характеристикой ферментативного потенциала почв является суммарная величина активности на вес столбика почвы с площадью поверхности 1 см2. Оказалось, что исследованные почвы различаются и по суммарной активпости протеазы и уреазы. Особенно различен ферментативный потенциал дерновой и сильноподзолистой почв, первая из которых содержит в 31 раз больше уреазы, в 6 раз протеазы и в 11 раз больше азота (см. табл. 5).
Интенсивность накопления аминокислот выше всего также в дерновой почве под лугом (см. табл. 4). В дерново-подзолистых поч- вад под лиственными породами суммарная активность протеазы в 2 раза, а уреазы в 7 раз выше, чем в сильноподзолистой почве под ельником зеленомошным (см. табл. 5). Ферменты в этой последней почве локализованы в основном в подстилочных слоях, обладающих высокой активностью на грамм образца, но общий объем потенциальных ферментативных превращений у них ниже, так как масса подстилки в несколько раз меньше массы гумусово-аккумулятивного горизонта, которого эта почва лишена. Выявление достоверных различий между этими почвами по интенсивности накопления аминокислот затруднено из-за высокой пространственной и сезонной изменчивости этого параметра.
Чтобы оценить вклад отдельных факторов и свойств почвы в создание ее ферментативного фонда, данные по активности протеазы за несколько лет были обработаны информационным методом (Карпа- чевский, 1977). Оценивалась связь уровня активности протеазы с (содержанием общего углерода, влажностью почв, генетическими горизонтами и типом растительности. Уровень активности протеазы разбивали на 8 групп, содержание углерода — на 5, влажность — на 9 групп. Ниже дана оценка связи между активностью протеазы на грамм почвы и характерующими биогеоценоз параметрами (одинарные каналы связи).
По степени убывания влияния на уровень протеазы эти параметры располагаются в следующий ряд: содержание углерода, влажность, генетический горизонт и, наконец, растительность, что подтверждает основные положения, изложенные выше.
Итак, показана возможность выявления специфических особенностей естественных почв, сформированных под разными фитоценозами, по уровню и характеру распределения в них каталитически активных метаболитов микроорганизмов — ферментов азотного обмена.
Далее была предпринята попытка выявить различие в биологическом режиме целинных и окультуренных почв, оценивая его по уровню активности протеазы, интенсивности накопления свободных аминокислот и по интенсивности «дыхания». Все эти процессы тесно связаны с численностью, составом и функциональной активностью живого населения почвы.
По мнению некоторых авторов, протеаза почв имеет в основном микробное происхождение (Петерсон, 1961; Купревич, 1968). Источником же свободных аминокислот и углекислого газа в почве, кроме почвенных организмов, служат и корневые выделения растений.
Изучались целинные под лесом и окультуренные почвы, расположенные в непосредственной близости от своих естественных аналогов па одинаковом элементе геоморфологического рельефа. Исследования проводили в двух районах Московской обл.: Подольском (Малинское лесничество) и Солнечногорском (агробиостанция «Чашниково»). В этой работе приводятся данные лишь за 1979 и 1981 гг. Сравнение почв по активности протеазы в пахотном и гумусовоаккумулятивном горизонтах обнаруживает существенные различия между окультуренными и целинными почвами, последние из которых содержат значительно больше углерода и азота. Активность протеолитических ферментов в гумусово-аккумулятивном горизонте целинных почв, сформированных как под хвойными, так и под лиственными породами, в несколько раз выше (в 2,2—4,5), чем в пахотном горизонте окультуренных почв ( 2, а, б).
Различия в их активности, как правило, достоверны при высоких уровнях значимости. Лишь в одном случае, когда активность протеазы в почве под березняком волосис- тоосоковом резко снизилась в конце августа 1979 г., она достигла уровня активности протеазы в залежной почве. Сезонные колебания активности протеазы более контрастны в целинной почве, что косвенно говорит о более резких колебаниях микробиологической активности этих почв. Важным обстоятельством является тот факт, что амплитуда и направленность сезонных изменений активности протеазы в целинных и окультуренных почвах порой не совпадают, что указывает на необходимость режимных наблюдений при оценке и выявлении специфики ферментативного потенциала почв.
Таким образом, уровень активности протеолитических ферментов устойчиво диагностирует потенциальные возможности катализа процессов азотного метаболизма целинных и окультуренных дерново- подзолистых почв. В связи с этим стоит вопрос об оценке биологической активности обследуемых почв с использованием полевых методов: по интенсивности накопления свободных аминокислот на аппликационных полотнах и по выделению углекислого газа или по интенсивности «дыхания».
Судя по данным табл., интенсивность накопления аминокислот крайне неоднородна, коэффициент пространственного варьирования (F) порой достигает 93%. Значительны и сезонные колебания, особенно велики они в окультуренной почве в год с большим дефицитом влаги. Так, в июне, когда запас влаги в почве был еще достаточен, ца полотнах за 30 дней накапливалось до 0,97 мг аминокислот на 1 г ткани, а в августе интенсивность накопления снизилась в 10 раз. В почве целинного участка за этот же период она почти не изменилась. В периоды с достаточным увлажнением в целинных и окультуренных почвах накопление аминокислот протекает примерно с одинаковой скоростью, особенно в верхнем 7-сантиметровом слое, который в целинной почве в наибольшей мере обогащен органическим веществом. Важно отметить, что в целинных почвах интенсивность .накопления аминокислот с глубиной резко снижается, что свойственно также и активности ферментов. В окультуренных же почвах в 25-сантиметровом слое процесс накопления аминокислот протекает с одинаковой интенсивностью.
Таким образом, замечена некоторая аналогия между изменением интенсивности накопления свободных аминокислот и активностью ферментов азотного обмена в целинных и окультуренных почвах. Однако освоение дерново-подзолистых почв сопровождается более существенным и устойчивым изменением уровня протеолитических ферментов.
Напряженность биологических процессов в почве
Учитывая данные о том, что суммарным показателем напряженности биологических процессов в почве является интенсивность выделения углекислоты (Мина, 1957), и данные о сопряженной связи активности ферментов и дыхания в некоторых типах почв (Попова, 1970), важно было выявить характер изменения и соотношение этих двух параметров внутри дерново-подзолистых почв и оценить их диагностическую ценность в изменении биологических свойств почвы под влиянием антропогенного воздействия.
Интенсивность дыхания целинных и окультуренных почв определяли абсорбционным методом (Карпачевский, 1977) на тех же пробных площадках, что и активность ферментов и интенсивность накопления свободных аминокислот. Показано, что интенсивность выделения углекислого газа с поверхности почв в значительной мере определяется гидротермическими условиями вегетационного периода и года (табл. 6). На всех почвах обнаруживались достоверные сезонные изменения интенсивности дыхания, величина которой в июне 1979 г. была максимальной в целинной почве и на залежи, достигая почти 4 кг/га за 1 ч, а к концу августа она снижалась примерно в 2 раза. Велик и коэффициент пространственного варьирования этого параметра. На фоне пространственного и временного изменения интенсивности дыхания различия между целинными и окультуренными почвами по этому показателю выявляются очень слабо и лишь в отдельные периоды наблюдений (июнь 1979 г. и август 1981 г.), в то время как по активности протеазы они различаются в 2—3 раза ( 3, а, б). Видимо, содержание гумуса, азота и другие агрохимические свойства дерново-подзолистых почв оказывают большее влияние на становление в них определенного ферментативного уровня, чем на интенсивность дыхания, величина которой и ее изменения определяются иными факторами.
Выводы
В профиле дерново-подзолистых почв, сформированных под естественной лесной растительностью, активность ферментов азотного обмена находится в корреляционной связи с содержанием углерода, азота, интенсивностью накопления свободных аминокислот на аппликационных полотнах и с влажностью почв.
В почвах, сформированных под разными естественными фитоценозами, выявлены специфические различия активности ферментов, отнесенной к грамму образца, суммарной активности в расчете на вес горизонта и столбика почвы с основанием в 1 см2, соотношения содержания ферментов в разных генетических горизонтах.
Определена временная и пространственная изменчивость уровня протеазы, уреазы, интенсивности накопления свободных аминокислот и выделения углекислого газа в целинных и окультуренных почвах.
Оценено влияние окультуривания дерново-подзолистых почв на активность протеазы, интенсивность накопления свободных аминокислот и выделения углекислого газа. Показано, что под влиянием окультуривания наиболее устойчиво изменяется ферментативный потенциал почв.
ЛИТЕРАТУРА
Аринушкина Е. В. Руководство по химическому анализу почв. М.: Изд-во МГУ, 1961. 491 с. Вайчис М., Данусявичус Ю. Влияние березы на свойства почвы под сосновым насаждением.—Почвоведение, 1978, № 1, с. 113—123. Галетян А. Ш. Ферментативная активность почв Армении.— Тр. НИИ почвоведения и агрохимии. Ереван: Айастан, 1974. 275 с. Гелъцер Ю. Г., Комовникова Т. Н. Биологическая активность лесных почв.— В кн.: Генезис и экология почв Центрально-лесного государственного заповедника. М.: Наука, 1979, с. 172—196 Дмитриев Е. А., Самсонова В. П. О пространственном варьировании содержания обменного кальция в дерново-подзолистой почве под ельником.— Науч. докл. высш. шк. Биол. науки. Сер. почвоведение, 1975, № 11, с. 112—118. Ивлева С. Н., Щербакова Т. А., Капитула А. Н. Биохимические процессы в дерново-подзолистых лёссовидно-суглинистых почвах под различными типами ельников Белоруссии.— В кн.: Тез. докл. VI делегат, съезда Всесоюз. о-ва почвоведов. Тбилиси, 1981, ч. 1, с. 155—156. Карпачевский J1. О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во МГУ, 1977. 312 с. Карпачевский Л. О., Киселева Н. КПопова С. И. Пестрота почвенного покрова под широколиственно-еловым лесом.— Почвоведение, 1968, № 1, с. 10—24. Колоскова А. В., Мурзатова С. Г. Формы азота и активность ферментов при азотном обмене в некоторых почвах Татарии.— Почвоведение, 1978, № 5, с. 58—65. Купревич В. Ф. Первые итоги исследований по ферментам почв.— В кн.: Сб. докл. симпоз. по ферментам почв. Минск, 1968, с. 3—10. Мина В. Н. Биологическая активность лесных почв и ее зависимость от физико- географических условий и состава насаждений.— Почвоведение, 1957, № 10, с. 73—79. Петерсон Н. В. Источники обогащения почв ферментами.— Мнкробиол. журн., 1961, т. 23, вып. 6, с. 5—11. Попова Э. П. О связи интенсивности дыхания почв и активности некоторых ферментов.— В кн.: Пути повышения урожайности сельскохозяйственных культур. Красноярск, 1970, с. 83—89. Раськова Н. В. Протеолитическая активность почв под естественными фитоце- нозами.— В кн.: Структурно-функциональные особенности естественных и искусственных биогеоценозов. Днепропетровск, 1978, с. 120—121. Раськова Н. В., Звягинцев Д. Г. Активность каталазы в почвах под широколиственно-еловым лесом и лугом.— Почвоведение, 1981, № 6, с. 76—82. Раськова Н. ВЗвягинцев Л. Г., Краснова М. Г. Активность гидролаз и оксидо- редуктаз в дерново-подзолистой почве с разным уровнем окультуренности.— Почвоведение, 1977, № 12, с. 124—129. Смолъянинов И. И., Рябуха Е. В., Чони Л. ИЗарудная Ю. А. Протеолитическая активность лесных почв в связи с интенсивностью биологического круговорота веществ в различных лесорастительных условиях.— В кн.: Лесоводство и агролесомелиорация: Респ. межвед. темат. науч. сб. Киев, 1968, вып. 15, с. 65—73. Хазиев Ф. XАгафарова Я. М. Протеолитическая активность черноземов Западного Предуралья.— Науч. докл. высш. шк. Биол. науки, 1975, № 5, с. 130— 135. Чернявский Я. В. Протеолитическая активность и восстановительная способность бурых горно-лесных почв и подстилок в буково-пихто-еловых лесах Украинских Карпат.— В кн.: Лесоводство и агролесомелиорация: Респ. межвед. темат. сб. Киев, 1976, вып. 46, с. 74—78. Щербакова Т. А. Роль ферментов в процессах трансформации поступающего в почву органического вещества.— В кн.: Экологические условия и ферментативная активность почв. Уфа, 1979, с. 59—77. Boss D. /. Protease aktivity and its relationship to nitrogen mineralisation in some soils under pasture and tussock grassland.— N. Z. J. Sci., 1977, vol. 20, N 2, p. 179—185. |











Азотный обмен в почвах
К содержанию книги: Почвенные организмы как компоненты биогеоценоза