|
|
Граница мела и палеогена на Малом Балхане и в Западном Копетдаге |
|
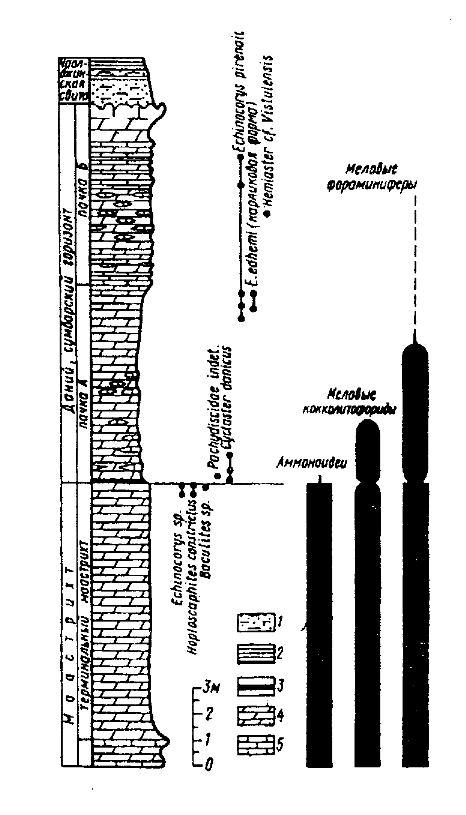
Пограничные отложения мела и палеогена в западной части Туркмении широко распространены, хорошо обнажены и довольно полно изучены. Нами в 1983 и 1985 гг. исследованы два района развития пограничных толщ - в долине р. Сумбар (Копетдаг) и на Малом Балхане (Алексеев и др., 1988). В первом районе наиболее интересен разрез СМ4, расположенный на северном крыле Сумбарской синклинали в 6 км от пос. Каракала на склоне горы Исак, примерно в 100 м восточнее шоссейной дороги на Кызыл-Арват. Терминальный маастрихт (общей мощностью 10 м) представлен голубовато-серыми мергелями, содержащими вверху довольно редкие панцири морских ежей рода Echinocorys (опр. М.М.Москвина) и многочисленные ядра аммонитов Hoploscaphites constrictus (J.Sow.), Baculites sp. Ядра аммонитов встречаются вплоть до кровли маастрихта и присутствуют даже в верхних 5 см меловых отложений (рис.9).
Рис. 9. Распределение основных групп ископаемых в разрезе СМ4 на р. Сумбар (Туркмения) В основании вышележащей сумбарской свиты (или горизонта, см. Калугин, Крымус, 1963), сложенной мергелями с линзовидными прослоями известняков (15 м) и принадлежащей датскому ярусу, наблюдается прослой коричневой и красновато-серой глины, обогащенной в базальных частях гипсом. Мощность этого слоя около 6 см. Его нижняя граница резкая, слегка волнистая, однако иногда на ней встречаются карманообразные углубления шириной до 15 см и глубиной 5-7 см, выполненные желто-бурым железистым материалом с большим количеством гипса. Коричневая глина вверх довольно резко переходит в зеленовато-серую известковистую глину (мощность последней 5 см). Суммарно коричневая и зеленая глина составляют 11 см и являются ПГГ. Пограничная глина постепенно переходит в зеленовато-серые мергели, которые незаметно сменяются глинистыми известняками с прослоями известняков. В мергелях содержатся довольно многочисленные панцири морских ежей Cyclaster danicus Schlьter, Pseudogibbaster sp. Здесь же (инт. +22 - +25 см над подошвой ПГГ) обнаружен единственный отпечаток аммонита с частично ожелезненным фрагмоконом (рис.), который не может быть определен точнее чем Pachydiscidae gen. indet. Этот аммонит был обнаружен в лаборатории при просмотре пробы, отобранной на микрофауну. Повторные поиски в 1985 и 1987 гг. на этом уровне с удалением больших объемов породы (несколько кубометров) положительных результатов не дали. Максимально высокая находка Cyclaster danicus сделана в одном метре от подошвы сумбарской свиты. Начиная с середины последней (6,5 от основания), панцири морских ежей вновь становятся довольно многочисленными. Среди них определены Echinocorys pyrenaica Seunes (инт. 6,5-12,7 м), E. edhemi Boehm (карликовые формы, инт. 7,0-7,5 м), Hemiaster cf. vistulensis Kongiel (10,5 м выше основания). Все эти виды морских ежей типичны для датского яруса Западной Европы, Северного Кавказа и Закаспия. Породы содержат обильные раковины фораминифер, но они обычно плохой сохранности в связи с погружением этих толщ на большую глубину. Распределение фораминифер в разрезе СМ4 изучено путем подсчета во фракции >0,05 мм. Мергели кровли маастрихта содержат довольно многочисленные раковины фораминифер (2500-3500 экз/г), среди которых планктонные формы составляют 76-89%. Планктонные фораминиферы характеризуются сравнительной бедностью таксономического состава и представлены Globotruncana arca Cushman, Globotruncanita stuarti (Lapp.), Globotruncanella petaloidea (Gand.), Pseudotextularia elegans (Rzehak), а также мелкими гетерогелицидами, которые доминируют среди планктона. Среди бентосных форм следует упомянуть Bolivinoides draco Marsson и Stensioeina pommerana Brotzen. Много кальцисферулид. Коричневая глина пограничного глинистого горизонта крайне бедна остатками фораминифер, особенно в ее нижней части. Так, в обр. СМ4/2 (0-1 см выше основания ПГГ) из 8 г. породы выделено лишь 12 раковин фораминифер Chiloguembelina sp. (2 экз.), Cibicides sp. (1 экз.), Globorotalites sp. (1 экз.), гладкие и ребристые прямые раковины нодозариид (8 экз.). Выше (обр. СМ4/3, 1-2 см) разнообразие комплекса возрастает - появляются особенно многочисленные Tappanina selmensis (Cushman) (до 20-30%), а также Globotruncanella petaloidea (Gandolf.), Globotruncanita stuarti, Clavulinoides kjrendagensis Moroz., Bolivina sp. Планктонные фораминиферы на этом уровне составляют 39%, агглютинирующий бентос - не более 4,5 %. В образцах СМ4/4 и СМ4/5 содержится заметное количество раковин Guembelitria cretacea Cushman (2,2-2,4%). Одновременно растет общее количество раковин фораминифер (от 1,5 экз/г в обр. СМ4/2 до 632 экз/г в обр. СМ4/5) и доля планктонных форм (до 48%). В нижней части зеленой глины, где много гипса, фораминиферовое число вновь падает до 104 экз/г, при этом соотношение планктона и бентоса не изменяется. В вышележащих мергелях общий облик комплекса фораминифер становится очень близким к маастрихтскому (значительную роль в нем играют глоботрунканиды, Pseudotextularia elegans, Bolivinoides draco, Stensioeina pommerana и другие меловые виды). Аналогичный облик фораминиферового комплекса сохраняется на протяжении почти всей остальной части сумбарской свиты до гиатуса в ее кровле. В 3 м от основания ПГГ встречены первые единичные палеоценовые планктонные фораминиферы Praemurica taurica (Moroz.), что указывает на зону Р1а (Berggren et al., 1995). Более полно распределение фораминифер непосредственно на К/Т границе изучено в соседнем разрезе СМ5, который расположен в 150 м западнее разреза СМ4 (Катана, Алексеев, 1991). Литологически этот разрез идентичен разрезу СМ4, но породы менее выветрелы. Кровля маастрихта сложена однородными мергелями, в которых присутствуют аммониты Hoploscaphites constrictus ( J. Sow.) (в 15 и 5 см ниже границы). В основании дания ПГГ включает шоколадного цвета глину с тонкими прослоечками гипса (0-2,5 см), бурую глину (2,5-4,5 см), переход к зеленой глине (4,5-6 см), зеленую глину (6-15 см) и переход к мергелю (15-20 см). Выше залегают обычные мергели, в которых на уровне 20-40 см от подошвы ПГГ встречены многочисленные панцири датских морских ежей Cyclaster danicus Schlьter. Опробованы верхние 1,5 м мергелей маастрихта, причем их последние 20 см охарактеризованы 5 образцами, и нижние 0,4 м дания. В ПГГ образцы отбирались бороздой с шагом по мощности 2-4 см, а выше - каждые 5 см. Всего проанализировано 18 образцов. Подсчет фораминифер производился раздельно по двум фракциям 50-160 и >160 мкм. В маастрихте содержание фораминифер высокое - обычно свыше 1000 экз/г, хотя отмечается некоторая тенденция к понижению вверх по разрезу. В ПГГ, за исключением базальных 2,5 см, содержание фораминифер стабильно низкое (около 200 экз/г), но выше чем в разрезе СМ5. Оно не изменяется при переходе от ПГГ к мергелям. Резкий рост до маастрихтского уровня отмечен в обр. 7 - 9 (25-40 см). Значительное падение фораминиферового числа в нижней части дания по сравнению с маастрихтом было отмечено В.Г.Морозовой и др. (1967) в сравнительно недалеко расположенном разрезе Кызылчешме. Среди фораминифер доминируют планктонные виды, которые в маастрихте составляют не менее 70% и около 50% в дании, однако подавляющую роль среди них играют гетерогелициды (обычно 75-80% от общего числа планктонных форм). Небольшой пик планктона отмечен вблизи кровли маастрихта, затем следует постепенное падение, достигающее минимума (39%) на уровне +20-25 см. Содержание бентосных агглютинирующих фораминифер крайне не велико (обычно доли процента) и не испытывает заметных колебаний. Не отмечается его рост и в ПГГ, который можно было бы объяснить растворением раковин секреционных фораминифер. Наиболее заметные изменения комплексов фораминифер связаны с резким увеличением численности Tappanina selmensis (Cushman), которое начинается в последних 3 см маастрихта (14,7%) и достигает максимума в ПГГ и перекрывающих мергелях (20-30%). В ПГГ отмечено массовое развитие Guembelitria cretacea Cushman (до 21%). Еще один пик доли этого вида отмечен на уровне +35-40 см. Доминирование этих двух видов-оппортунистов, переживающих рубеж мела и палеогена повсеместно типично для зоны P0 основания датского яруса.
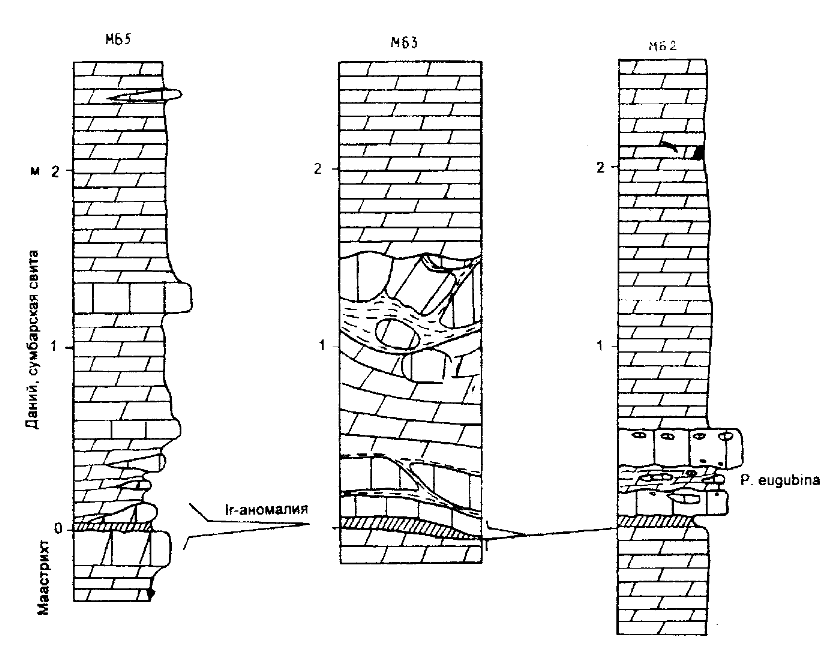
Рис. 10. Цунамигенная (?) единица в подошве датского яруса на Малом Балхане В разрезах Малого Балхана интерес представляют признаки цунамигенных отложений (рис.10). В наиболее южном разрезе МБ5 переход от маастрихта к данию выглядит также как и в разрезе СМ5. Однако в 1,5 км северо-восточнее в разрезе МБ2 над редуцированным ПГГ (до 5 см) с очень слабой иридиевой аномалией залегает пласт массивного прочного известняка (15 см), содержащего редкие линзовидные литокласты известняка аналогичной окраски и структуры. Подошва и кровля известняка резкие и волнистые, он перекрыт рыхлым глинистым мергелем (12-16 см) с линзовидными включениями известняка. Мергель наряду с маастрихтскими фораминиферами содержит Parvularugoglobigerina eugubina зоны Р1а нижнего дания. Завершается эта пачка массивным пластом известняка (20-25 см), в подошве и кровле которого заключены овальной формы известняковые интракласты и единичные галечки розоватого известняка. Кровля пласта очень резкая. В 200-300 м северо-восточнее в разрезе МБ3 непосредственно выше ПГГ (до 7 см) с хорошо выраженной иридиевой аномалией (Назаров и др., 1988) залегают два пласта известняка, верхний из которых имеет линзовидное строение. Они разделены глинистыми прослойками. Мощность пластов 10-20 см. На известняках лежат неяснослоистые мергели (0,4-0,6 м), в верхней части которых обособляются часто с размытыми границами включения известняков. Завершается разрез глинистым мергелем (0,4 м) с включениями разно ориентированных и неправильной формы, но с оглаженными краями известняковых отдельностей размером до 0,3 х 0,5 м. По четкой границе известняки перекрываются мергелями. Образование известняковой пачки непосредственно выше глинистого горизонта с иридиевой аномалией несомненно связано с перемещением слабо консолидированных осадков в виде обломочных потоков и оползней вниз по склону бассейна, мелководье которого располагалось к северу от Малого Балхана в платформенной части Туркмении. Спусковым механизмом могло быть цунами или кратковременное эвстатическое падение уровня моря. Скорее всего, малобалханская обломочная единица и цунамиты Мексиканского залива не вполне одновозрастны, так как первая залегает не ниже, а выше ПГГ с иридиевой аномалией. В Южной Голландии перемытые отложения маастрихта также содержат нанопланктон зоны NP1 с Biantholitus sparsus, мелкими Neobiscutum и Criuciplacolithus primus (Romein et al., 1996). В Бразилии брекчии имеют близкий возраст - верхняя часть зоны Р1а (Kotsoukos, 1995).
|
К содержанию: Алексеев А.С. "Массовые вымирания в фанерозое"
Смотрите также:
Палеонтология Дрейф материков и климаты Земли Палеоклиматология
Геохронологическая таблица Геологическая шкала. Абсолютный и относительный возраст горных пород
Причины вымирания динозавров Гипотезы вымирания динозавровой фауны
Причины вымирания древних фаун Почему вымерли динозавры Фанерозой
Криптозой и фанерозой в истории Земли. Биогенные породы.
Криптозон и фанерозой. Климаты позднего докембрия и палеозоя.
Как запомнить геологические эпохи - каждый отличный студент...
Изменение солености океана в фанерозое
Периодизация истории земли. геохронологическая... Метеориты и кометы