|
|
Меловой период Мезозойской эры. Белемниты и аммониты. Возникновение цветковых покрытосеменных растений |
Мезозойская эра заканчивается меловым периодом, пробравшимся около 70 млн. лет. Он знаменовал собой качественно новый этап в развитии ландшафтно-климатических областей и органического мира Земли. В течение этого периода тектоническая жизнь планеты оставалась активной.
В течение мелового периода продолжались киммерийские горообразовательные движения. В начале мелового периода в результате этих движений были созданы горно-складчатые сооружения на востоке Евразии. Складчатые пояса располагались на огромной территории, охватывающей Верхоянье, Чукотку, Камчатку, Дальний Восток и восток Китая. Дополнительные складкообразовательные движения на границе раннего и позднего мела (так называемая австрийская фаза складчатости) усилили контрастность этих участков земной коры. В это время, как и ранее, наряду с подводными вулканическими излияниями осуществлялось внедрение гранитных интрузий.
Движения, обусловленные австрийской фазой складчатости, наиболее сильно проявились в Восточной Азии и Кордильерах и менее интенсивно в современном Альпийско-Гималайском поясе (Восточные Альпы, Карпаты, Кавказ, Памир). В конце мелового периода имела место довольно активная ларамийская орогеническая фаза. Складчатые движения охватили окраины Тихого океана. На значительную высоту оказались приподняты Анды, Кордильеры, горные сооружения Антарктиды, Чукотки и Камчатки.
Продолжалось раздвижение гондванских континентов. Северная Америка отделялась от Африки и Евразии. Эти активные раздвижения сопровождались формированием новых морей и океанов. Так, например, Атлантический океан приобрел близкие к современным очертания, возникли Бискайский залив и Гренландское море. В это время был заложен рифт Красного моря и началось раскрытие Индийского океана, а Индостан и Мадагаскар отделились от Африки. Индостанский континент совершил длительное путешествие, исчисляемое тысячами километров, пока в середине палеогена он не столкнулся с Азиатским континентом. В результате такого столкновения были образованы Гималаи.
Началу мелового периода был свойствен регрессивный этап развития, но в середине раннего мела ему на смену пришел трансгрессивный этап. Позднемеловая морская трансгрессия была одной из самых величайших трансгрессий на Земле. В это время многие континенты оказались покрытыми морем. Море занимало почти 40% территории современной Евразии и 35% территории Северной Америки, около 20-25% территории Африки и Австралии и 15% территории Южной Америки.
В тесной зависимости от тектонических движений формировался рельеф земной поверхности. Наиболее контрастным и возвышенным он был в местах столкновения литосферных плит между собой. В результате поднятии и складчатости были образованы многие горные массивы. Области платформенной активизации также обладали возвышенным и горным рельефом.
Так же как и в настоящее время, горные сооружения и глубоководные океанические впадины занимали на земной поверхности весьма незначительную площадь. Основная же роль принадлежала выровненным шельфовым участкам, неглубоким эпиконтинентальным морям, приморским озерно-дельтовым и внутриконтинентальным озерно-аллювиальным низменностям и равнинам, возвышенным равнинам с увалисто-холмистыми водоразделами и выровненным плато. На них, а также на плоских водоразделах в благоприятных климатических условиях формировались коры выветривания. Некоторые более древние горные сооружения (герциниды, каледониды) в эпохи активизации испытали поднятия, однако амплитуда высот была не очень значительной и вряд ли превышала 1000 м.
В течение мелового периода развитие организмов и их расселение определялись природными условиями. Значительные изменения в распределении ландшафтно-климатических зон в первую очередь отразились на наземной флоре. В середине мелового периода осуществлялось широкое расселение многих представителей высшей флоры - покрытосеменных растений. Еще в начале мелового периода их роль в растительном покрове была невелика, но уже в середине позднемеловой эпохи они заняли главенствующее положение.
По сравнению с поздней юрой в раннем мелу произошло изменение морфоанатомического строения гинкговых. В то время как юрские гинкговые обладали сильно расчлененной листовой пластинкой, раннемеловые формы имели уже цельную пластину. Это свидетельствует не столько о более высокой ступени развития, сколько об их приспособлении к сезонным, а возможно, и к суточным колебаниям температур.
Многие древесные сообщества хвойных и гинкговых, особенно те, которые росли в высоких и средних широтах, обладали ясно выраженными годичными кольцами. Это свидетельствует о существовании в зоне хвойно-гинкговых лесов относительно прохладных условий.
В средних широтах располагалась зона цикадофитово-беннеттитовых лесов. Она отделялась от зоны хвойно-гинкговых лесов довольно узкой полосой развития смешанных цикадофитово-хвойно-гинкговых лесов. В Евразии к этой зоне относились крайний север Европы, Северный Урал, юг Западной Сибири и Приморье.
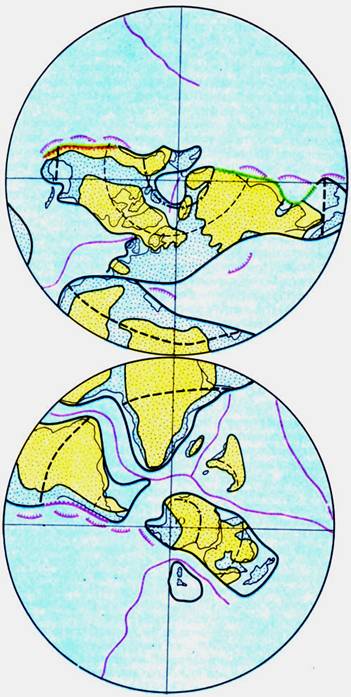
Положение литосферных плит и срединно-океанических хребтов в позднемеловой эпохе (по А. М. Городницкому и Л. П. Зоненшайну)
 Наивысшего расцвета и
разнообразия достигла флора в областях с тропическим влажным климатом. Склоны
возвышенностей и низменности были покрыты густым пологом леса, весьма сходного
с современными экваториальными лесами. Особенно большого разнообразия достигли
цикадофитовые и многочисленные древовидные папоротники. В сторону областей с засушливыми
условиями многие влаголюбивые формы исчезали, лесной покров редел и превращался
в ксерофильное редколесье.
Наивысшего расцвета и
разнообразия достигла флора в областях с тропическим влажным климатом. Склоны
возвышенностей и низменности были покрыты густым пологом леса, весьма сходного
с современными экваториальными лесами. Особенно большого разнообразия достигли
цикадофитовые и многочисленные древовидные папоротники. В сторону областей с засушливыми
условиями многие влаголюбивые формы исчезали, лесной покров редел и превращался
в ксерофильное редколесье.
Лесная растительность тропического типа произрастала в Южной Евразии, на юге Северной Африки, юге Гренландии, а также в Африке, Австралии и Южной Америке.
В начале раннемеловой эпохи в лесах, состоящих из голосеменных форм, появились первые представители покрытосеменных растений. Уже в конце раннего мела их роль резко увеличилась. Сильно уменьшился видовой состав голосеменных, несмотря на существование довольно благоприятных условий. Появились и сравнительно быстро распространились по всему земному шару магнолии, лавровые, платаны, эвкалипты и дубы. Среди хвойных господствующее положение заняли сосновые и таксодиевые. Эти две группы растений по условиям произрастания сильно отличались друг от друга. Сосновые леса, хорошо приспособленные к интенсивному солнечному облучению и засушливым условиям, произрастали на открытых возвышенных участках. Таксодиевые же, являющиеся тенелюбивыми растениями, предпочитали области с сильным увлажнением. Поэтому заросли таксодиевых располагались в низменных, обильно увлажненных местах, главным образом на низких поймах. В конце раннего мела постепенно исчезли многие мезозойские формы (подозамитовые и саговые) и сильно обновился состав папоротников, особенно среди травянистых форм.
В раннемеловую эпоху наземные формы животных достигли большого разнообразия. На суше по-прежнему господствовали хорошо приспособленные к обитанию в лесных и саванных ландшафтах рептилии. Наличие благоприятных условий для жизнедеятельности, обилие пищи и отсутствие врагов способствовали у них развитию гигантизма.
По сравнению с позднеюрской эпохой в раннем мелу число зауропод уменьшилось, господствующее положение заняли птиценогие динозавры - игуанодонты и утконосные динозавры. Господство двуногой динозавровой сухопутной фауны было связано с необходимостью обитания среди высокой растительности и периодическим пребыванием в воде. Эти крупные травоядные животные жили главным образом в болотистых лесах, расположенных на берегах крупных пресноводных водоемов. Не имея иных средств защиты, они скрывались в озерах, заводях и заливах от хищных динозавров - тарбозавров и тиранозавров. Так же как и позднеюрские предшественники растительноядных динозавров, игуанодонты имели на задних конечностях перепонки, что позволяло им беспрепятственно, несмотря на огромные размеры, передвигаться по влажным и топким грунтам.
Типично сухопутными формами были растительноядные четвероногие рептилии. Для защиты от хищников они имели тяжелые панцири с шипами, роговыми воротниками, колючими гребнями или толстую шкуру, покрытую мелкими, но плотными роговыми пластинками.
Обилие атмосферных вод и наличие неровностей рельефа способствовали возникновению пресных озер, в которых наряду с различными рыбами обитали довольно многочисленные пресноводные брюхоногие и двустворчатые моллюски. В озерах, зарастающих водной и болотной растительностью (и в связи с этим обедненных кислородом), моллюсковая фауна была угнетенной и весьма малочисленной.
В раннемеловую эпоху среди морской фауны беспозвоночных осуществилась дифференциация по климатическому признаку. В приполярных морях распространялась бореальная фауна, а в низких широтах - тропическая и экваториальная. Четкой границы между этими двумя довольно различными комплексами фауны практически не существовало. В морских бассейнах, занимавших промежуточное положение, обитали смешанные фауны, в составе которых в зависимости от температурных условий преобладали то тропические, то бореальные представители. Бореальная фауна характеризовалась присутствием эндемичных родов аммонитов, белемнитов и двустворчатых моллюсков. Тропические моря были населены богатой фауной беспозвоночных. Основную роль играли кораллы, морские ежи, аммониты, белемниты, брахиоподы, теплолюбивые представители планктонных фораминифер и наннопланктон.
На морском мелководье, там где воды не испытывали сильных изменений солености ввиду отсутствия вблизи речного стока, находились крупные отмели, на которых располагались колонии устриц и рудистов, барьерные и береговые рифы. В районах развития рифовых комплексов обитали брахиоподы, морские ежи, морские лилии и не дожившие до настоящего времени строматопоры. Обширные пространства занимали подводные луга. На некотором удалении от берега находилось основное местообитание белемнитов и аммонитов.
В конце раннего мела ареал обитания бореальной фауны сильно расширился и многие из типичных бореальных форм (например, двустворчатые моллюски рода ауцеллин или аммониты рода краспедитес) достигли тропических морей.
В течение раннемелового времени в результате изменения температурного режима происходило то расширение, то сужение климатических поясов. Существовавший в начале раннемеловой эпохи значительный дефицит влаги постепенно сменился обильным увлажнением в ее конце. Сильно увеличились площади c гумидным типом климата. Одновременно с этим произошло незначительное общее похолодание.
Температуры приповерхностных морских вод в экваториальном и тропическом поясах на протяжении большей части раннего мела были довольно высокими. Например, в морях Западной Европы в начале раннего мела они составляли 20-25°С. Примерно такие же температуры существовали в конце раннего мела в морях, расположенных на востоке Южной Африки, в Северной Австралии и Северной Америке.
В самом конце раннего мела произошло снижение температур на 2-3°, и это незамедлительно сказалось на составе фауны. Резко сократились ареалы кораллов и рудистов, появились не только организмы, которые могли существовать в различных температурных условиях, но и бореальные организмы, а также уменьшилось карбонатонакопление.
Экваториальный пояс охватывал север Южно-Американского континента, центральную часть Африки, юг Аравии, Индию и Индокитай. По обе стороны от него находились области с тропическим засушливым климатом. Северный аридный сектор охватывал юг США, Мексику, Центральную Америку, север Африки и Южную Евразию, а южный - Южную Америку, Африку и центральные области Австралии. В этих областях преобладали пустыни и полупустыни с ксерофильным редколесьем, весьма сходные с современными саваннами.
Понижение температурного режима в конце раннего мела произошло повсеместно. Так, снижение среднегодовых температур на 5-8°С по сравнению с началом раннемеловой эпохи установлено для морей, расположенных на территории Карпат, Крыма, Северного Кавказа, юга европейской части СССР, Средней Азии и Сибири.
Прохладные условия в течение раннемеловой эпохи существовали в пределах северных окраин Северо-Американского и Евразиатского материков, на юге Африки, Австралии, Южной Америки и в Антарктиде. На севере Евразии среднегодовые температуры в течение продолжительного времени колебались от 12 до 15°С, а в конце раннего мела они составляли 8-12°С. Похолодание привело к смещению климатической зональности, и в приполярных районах возник климат, сходный с современным умеренным климатом.
Как уже отмечалось выше, в конце раннего мела произошла существенная перестройка растительного покрова. Этот важнейший в истории Земли и биосферы факт до настоящего времени все еще не нашел полного объяснения. Неясны причины и места возникновения покрытосеменной растительности. По-видимому, в это время произошли достаточно сильные преобразования в природных условиях, которые способствовали широкому распространению покрытосеменной флоры. Последняя была лучше приспособлена к динамическим контрастам ландшафтно-климатических условий, которые можно объяснить изменением состава атмосферы, тектонической активностью планеты и, возможно, космическими причинами. В конце раннего мела продолжались активные перемещения континентов, сильно изменилась концентрация углекислого газа и кислорода в атмосфере, осуществилось глобальное понижение температур, появились сезонные колебания температур и влажности, уменьшилась облачность, а следовательно, усилился приток как яркого солнечного света, так и жесткой радиации. Очевидно, эти факторы и привели к изменению соотношения мало приспособленной к резким колебаниям температур голосеменной и споровой флоры и широкому распространению лучше приспособленной покрытосеменной.
В отличие от голосеменных покрытосеменные растения обладают большей фотосинтезирующей поверхностью, у них более совершенные водно-сосудистая и корневая системы и они могут лучше переносить сезонные колебания температур и влажности.
В позднемеловое время хвойно-широколиственные леса произрастали в высоких широтах северного и южного полушарий. В конце позднего мела появились и сильно расширили свои ареалы листопадные теплоумеренные формы - орех, бук и береза. Хвойные были представлены сосновыми (сосна, ель, пихта, тсуга, кедр) и таксодиевыми. Однако еще в течение длительного времени в приполярных районах присутствовали реликты не только раннемеловой, но и даже юрской флоры. Здесь располагались заросли из низкорослых форм цикадофитов, араукариевых и гинкговых. В средних широтах произрастали широколиственные леса - платаны, лавры, дубы, каштаны и клены. В субтропиках росли как широколиственные листопадные, так и вечнозеленые магнолии, лавры, пальмы, а также секвойи, эвкалипты, сосны; в подлеске располагались папоротники, гинкговые и цикадофитовые.
Во влажных тропиках и в приэкваториальной части произрастали дождевые экваториальные леса, вечнозеленая растительность которых была представлена главным образом пальмами, древовидными папоротниками, вечнозелеными дубами, магнолиями и т. д.
В засушливых ландшафтах ксерофильного редколесья и сухих опустыненных саванн господствовали узко- и мелколистные формы растительности. Рощи и небольшие леса встречались лишь в виде оазисов и вдоль речных долин и образовывали галерейные леса.
Среди наземной фауны позвоночных в течение позднего мела появились новые виды четвероногих растительноядных рептилий - анкилозавры, цератопсы, которые обладали пассивными средствами защиты от хищников (шипы, панцирь, роговой покров). Они уже не жили, как их предки, в прибрежно-морских или приозерных зарослях, а переместились в пределы саванного редколесья. В этих ландшафтах водились рогатые цератопсы, очень сходные и в экологическом, и в морфологическом отношении с появившимися в кайнозое носорогами. В саванных ландшафтах существовали утконосые динозавры - гадрозавры. Они перемещались на двух ногах и были очень тесно связаны с водной средой. Об этом свидетельствуют, во-первых, утиная морда с далеко отодвинутыми ноздрями и, во-вторых, наличие плавательных перепонок на ногах. Гадрозавры питались прибрежной болотной растительностью, обитали в широких речных руслах, дельтах рек и по образу жизни напоминали современных бегемотов.
Большого разнообразия в позднемеловое время достигли крокодилы, основным местообитанием которых являлись реки, пойменные озера и старицы. Условия открытой саванны с умеренно аридным климатом и сезонным, но обильным выпадением осадков благоприятствовали развитию динозавровой фауны. Динозавры обитали в многочисленных тихих водоемах, окруженных зарослями из влаголюбивых растений. В заболоченных тропических и субтропических лесах на внутриматериковых аллювиальных и приморских озерно-дельтовых низменностях с обильным увлажнением происходило резкое сокращение видового состава динозавров. В субтропиках это объяснялось не столько высокой влажностью, сколько умеренностью климата и почти сплошным развитием озерно-болотных ландшафтов.
Слабое распространение позвоночных в лесных ландшафтах было связано с неблагоприятными условиями их обитания, непроходимостью лесных зарослей и огромных заболоченных пространств, трудностью пропитания.
В течение позднего мела широкое развитие получили пресноводные моллюски с массивными раковинами. Большого разнообразия достигла фауна морских беспозвоночных, представленная головоногими моллюсками - брахиоподами, двустворчатыми и брюхоногими моллюсками, кораллами, фораминиферами, морскими ежами и лилиями. Распространение всех беспозвоночных определялось температурным режимом. Так, в приполярных районах и морях высоких широт обитал бореальный комплекс, состоящий из песчаных фораминифер и эндемичных видов аммонитов и белемнитов. В нем отсутствовали кораллы и многие моллюски с массивной раковиной. В тропических морях фауна характеризовалась большим разнообразием. Многочисленные кораллы и строматопоры участвовали в построении крупных рифовых массивов. В мелководных морях обитали аммониты, брахиоподы, крупные и мелкие фораминиферы, рудисты и крупные устрицы.
На протяжении позднемеловой эпохи на Земле существовали экваториальный, тропический, субтропический и умеренный пояса. Первые два в отличие от аналогов предшествующей эпохи захватывали более обширные территории. В тропическом поясе обитали теплолюбивые организмы, произрастали вечнозеленые дождевые леса и располагались саванны. Температурный режим повсеместно был довольно высоким. Максимально высокие температуры существовали в середине позднемеловой эпохи. В морях Западной Европы они колебались в пределах 19-25°С, но в конце позднего мела, особенно на границе с кайнозоем, за исключением Южной Европы, среднегодовая температура понизилась до 15°С.
Высокие среднегодовые температуры были характерны для Карпат, Крыма, Закавказья и юга Средней Азии, в течение позднемеловой эпохи они изменялись в пределах 18-27°С. Высокие температуры в течение позднемеловой эпохи существовали в морях, располагавшихся на территории США (20-26°С), на юге Канады (20-28°С) и севере Австралии (20-24°С).
В северном полушарии находились два аридных сектора с тропическим климатом. Один из них охватывал север Африки, Аравию, Восточное Средиземноморье, а другой простирался от Средней Азии через центральные районы Китая до современного побережья Тихого океана. В южном полушарии аридные секторы занимали запад Южной Америки, центральные районы Африки и Индокитай. Для этих областей были характерны пустынные и полупустынные ландшафты и слабое развитие гидрографической сети. В сторону районов с высоким увлажнением пустынные ландшафты сменялись ксерофильным редколесьем.
За пределами тропического пояса располагались районы с относительно низким температурным режимом. На севере европейской части территории СССР, на юге Западной и Восточной Сибири и на Дальнем Востоке среднегодовые температуры изменялись и составляли 14-18°С.
В местах максимального увлажнения, где находились обширные приморские озерно-дельтовые и внутриконтинентальные озерно-аллювиальные низменности и равнины, росли смешанные широколиственные леса с примесью хвойных и папоротников. Умеренный пояс в северном полушарии охватывал северо-восток Евразии и северо-запад Северной Америки, а в южном полушарии выделялся на юго-востоке Австралии и в Антарктиде. Со временем климат приобрел черты континентальности, и главным показателем этого являются резкие суточные и сезонные перепады температур. Среднегодовые температуры в умеренном поясе обычно составляли 10-14°С, но в отдельные промежутки времени они повышались и это приводило к изменению положения климатических границ.
К содержанию: Ясаманов Николай Александрович «Популярная палеогеография»
Смотрите также:
Историческое землеведение палеогеография... Палеогеография пустыни Гоби Палеонтология и палеогеография палеогеография
Последние добавления:
Кроманьонцы Холопы на Руси Кондараки - Описание Крыма Палеонтология позвоночных Занимательная минералогия