|
|
БИОЛОГИЯ ПОЧВ |
КРУГОВОРОТ СЕРЫ
|
Смотрите также:
Мейен - Из истории растительных династий
Биографии биологов, почвоведов
|
Сера — биогенный элемент, необходимый для жизни. В белках сера содержится в форме некоторых аминокислот (цистин, цистеин), входит в молекулы витаминов, коферментов, присутствует в растительных эфирных маслах. В растениях содержание серы колеблется от 0,2 до 1,8%, в организме человека — около 0,4%. Соединения серы в живой клетке участвуют в важных метаболических процессах (например, она входит в состав ферментов нитрогеназы и нитратредуктазы, ответственных за превращения азота — его фиксацию и восстановление).
С урожаем из почв ежегодно выносится от 10 до 80 кг/га серы в зависимости от биологических особенностей сельскохозяйственных растений и величины урожая. Так называемые безбалластные минеральные удобрения не содержат серы, и дефицит ее в почве с ростом урожаев постоянно увеличивается. Поэтому необходимы изучение серного режима почв, разработка методов диагностики дефицита серы и эффективного применения серосодержащих удобрений. В почвах сера претерпевает разнообразные превращения, переходя из неорганических соединений в органические и обратно. В почве в виде неорганических соединений сера бывает окисленной (сульфаты, политионаты), восстановленной (сульфиды) и редко молекулярной.
При разложении остатков животных, растений и микроорганизмов освобождаются серосодержащие аминокислоты, тиоспирты, тиофенолы, тио- эфиры, гетероциклические соединения (например, тиофен), в которых сера находится в восстановленном состоянии. В органических веществах сера обычно содержится в восстановленном состоянии в виде SH-групп. Есть органические соединения, содержащие серу в окисленной форме. Это сульфиновые и суль- фокислоты и их соли, применяемые в моющих средствах как поверхностно-активные вещества (ПАВ), но это неприродные соединения. Они могут попадать в почву со сточными водами.
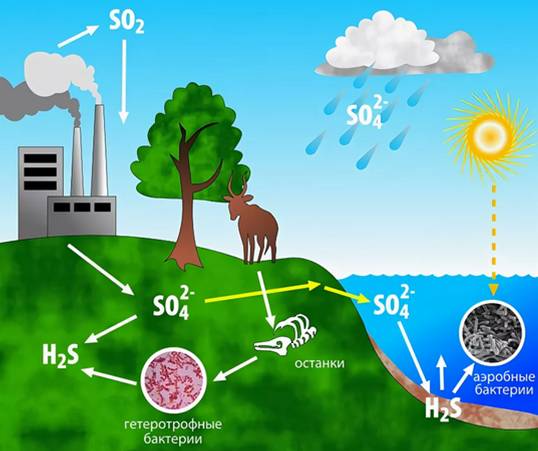
Цикл превращений серы сходен с циклом азота: он включает окислительные и восстановительные звенья, а также превращение серы без изменения ее валентности (84). В этих превращениях участвует много разнообразных групп микроорганизмов: аэробных и анаэробных, хемо- и фототрофов, истинных бактерий и архебактерий. В последние десятилетия было сделано много открытий в микробиологии цикла серы в связи с изучением архебактерий. Некоторые из них оказались активными участниками превращения соединений серы, способными осуществлять неизвестные ранее процессы.
Рассмотрим цикл серы, объединив в отдельные звенья процессы окислительного характера, восстановительные и процессы освобождения серы при разложении органических веществ.
Окисление серы и ее восстановленных неорганических и органических соединений происходит в аэробных и анаэробных условиях с участием разных групп микроорганизмов. В аэробных условиях окислительные процессы осуществляют хемоавтотрофные прокариоты — серные бесцветные (неокрашенные) бактерии и тионовые бактерии, термоацидофильные архебактерии, а также некоторые типичные гетеротрофные бактерии. В анаэробных процессах участвуют фототрофные серные пурпурные и зеленые бактерии, осуществляющие бескислородный фотосинтез.
Бесцветные серобактерии по строению сходны с цианобактериями. Основное их отличие от последних — отсутствие пигмента. По морфологии среди них различают (85) одноклеточные: крупные подвижные (Thiovulum) и неподвижные (Achromatium) бактерии; виды с мелкими спиралевидными или другой формы клетками (Thiospira, Thiobacterium); нитчатые многоклеточные неподвижные (Thiothrix) и скользящие бактерии (Beggiatoa, Thioploca). Общим признаком всех организмов этой группы служит способность откладывать внутриклеточно образующуюся при окислении H2S молекулярную серу. Механизм этого процесса и его физиологический смысл, возможно, различен у разных представителей и до конца еще не ясен.
С.Н. Виноградский впервые предположил возможность использования микроорганизмами энергии окисления неорганических соединений для синтеза органических веществ из С02 — хемосинтеза.
Четкие доказательства наличия хемосинтеза в чистых культурах бесцветных серобактерий были получены только в последнее время, например у Beggiatoa. В то же время известны случаи окисления сероводорода в результате гетеротрофной жизни некоторых бактерий из группы бесцветных серных за счет образования ими перекиси водорода при дыхании. Физиологический смысл этого процесса заключается в детоксикации среды путем разложения образующегося токсичного продукта метаболизма Н202 с участием сульфидов, которые окисляются до молекулярной серы.
Фотосинтезируюшие серные бактерии в отличие от бесцветных анаэробы. Наличие в их клетках пигментов бактериохлоро- филлов придает им красную или зеленую окраску, поэтому их делят соответственно на пурпурные (пор. RhodospiriHales) и зеленые (пор. Chlorobiales). Все они осуществляют на свету бескислородный фотосинтез, используя восстановленные соединения серы в качестве доноров электрона в анаэробных условиях.
Пурпурные серобактерии — грамотрицательные одноклеточные прокариоты разной формы и размеров, подвижные и неподвижные (см. 85). В клетках существует хорошо развитая система мембран — тилакоидов (внутрицитоплазматические разрастания мембраны), у некоторых есть газовые вакуоли. Окисляя H2S внутриклеточно, они временно откладывают в клетках молекулярную серу в виде капель, ограниченных белковой мембраной, а при дефиците сероводорода окисляют серу далее до серной кислоты. Представители: Thiospirillum и Chromatium — крупные подвижные бактерии; Thiodictyon, Thiocapsa— неподвижные, без жгутиков.
Зеленые серобактерии тоже одноклеточные, разной морфологии — от простых палочек до звездчатых клеток с простеками (нитевидные выросты клеток). Некоторые образуют цепочки или сетчатые структуры. Пигменты локализованы в хлоросомах. Зеленые серобактерии — более строгие анаэробы, чем пурпурные. Серу в клетках не накапливают, а выделяют ее наружу. Для большинства зеленых серобактерий установлена способность к азот- фиксации. Помимо одноклеточных зеленых серобактерий — Chlorobium, Prosthecochloris и других — известны представители зеленых скользящих бактерий с нитчатым строением (Chloronema), которые тоже способны к окислению H2S (см. 85). Встречаются в почвах рисовых полей и в болотах. Для их развития нужен свет и анаэробные условия.
Тионовые бактерии — хорошо изученная группа бактерий, так как они используются в гидрометаллургии для получения ценных металлов из бедных руд — хемолитоавтотрофы, использующие энергию окисления серы кислородом для процессов хемосинтеза. Эту группу составляют представители нескольких родов грамотрицательных эубактерий (Thiobacillus, Thiosphaera, Thiomi- crospira) и один род термоацидофильных архебактерий (Sulfolobus). Полное ферментативное окисление молекулярной серы тионо- выми бактериями приводит к образованию серной кислоты: S° S032" S042". Электроны поступают в дыхательную цепь. Кроме молекулярной серы и сульфида, тионовые бактерии могут окислять сероводород, тиосульфат, политионаты (S2062-, S4062"), тиоцианат (CNS~) и другие восстановленные или не до конца окисленные соединения серы. Образующаяся как промежуточный продукт окисления молекулярная сера всегда откладывается вне клеток. Некоторые тионовые бактерии в анаэробных условиях способны использовать вместо 02 нитраты, восстанавливая их до нитритов (Thiobacillus thioparus) или до N2 (Th. denitrificans). Для Sulfolobus и Thiobacillus ferrooxidans показана способность окислять и серу, и двухвалентное железо (Fe2+).
В природе бактерии, окисляющие серу, занимают специфические экологические ниши и встречаются в массовых количествах как в сильнокислых, так и в щелочных средах, где имеется сероводород или другие восстановленные соединения серы. Некоторые живут при высоких температурах в горячих серных источниках. Фототрофные серобактерии распространены главным образом в водоемах, хемотрофные тионовые — в почвах. Образуемая ими серная кислота подкисляет почву, что способствует переводу некоторых важных для растений элементов (фосфора, железа) в доступную форму. В то же время серная кислота может быть опасным агентом коррозии металлических труб и других сооружений в почве. Этот процесс подробно изучается, устанавливаются почвы, где он более возможен, составляются специальные картосхемы, разрабатываются меры по его предотвращению.
Тионовые бактерии используются в гидрометаллургии. Бактериальное выщелачивание металлов за счет окисления содержащихся в руде сульфидов позволяет вести добычу металла из бедного минерального сырья при минимальных энергозатратах. Разработано целое направление микробной гидрометаллургии (получение цветных металлов).
Восстановление сульфатов происходит в разных метаболических процессах. Сульфаты служат источником серы как для растений, так и для микроорганизмов. Их ассимиляция сопровождается восстановлением серы в процессах конструктивного метаболизма. Это ассимиляционная сульфатредукция сходная с соответствующим процессом поглощения и восстановления нитратов. Процесс этот универсален для всех организмов. Биологическое закрепление серы в микробных клетках называется также ее иммобилизацией.
В анаэробных условиях сульфаты восстанавливаются до сероводорода специфическими бактериями из группы облигатных анаэробов. Это узкоспециализированные сульфатредуцирующие бактерии, использующие сульфаты в качестве окислителя органических соединений в процессе анаэробного дыхания — дисси- миляционной сульфатредукции (сравни с денитрификацией). Они были открыты М. Бейеринком. Некоторые виды могут окислять свободный водород, выступая как хемолитоавтотрофы или как хемолитогетеротрофы. Для последних донором электрона служит Н2, а источником углерода — органические вещества, например лактат. У сульфатредуцируюших бактерий перенос электрона происходит через цитохромную цепь, как и у аэробных бактерий, с образованием АТФ. Возбудители процесса сульфатредук- ции относятся к 10 разным родам строго анаэробных бактерий (около 50 видов): Desulfovibrio — подвижные изогнутые палочки, не образующие спор, Desulfotomaculum — споровые палочки. Среди представителей первого рода есть галофилы, среди вторых — термофилы. В последние годы выделено много новых сульфатреду- цирующих бактерий: Desulfonema, Desulfosarcina, Desulfobacter, Desulfococcus. Два последних рода — кокковые бактерии, Desulfonema — крупные трихомные микроорганизмы со скользящим движением. Распространены сульфатредуцирующие бактерии во всех почвах, но особенно много их в почвах с режимом, приводящим к длительному анаэробиозу, например в затопляемых почвах рисовых полей, а также в болотах, илах, лиманных грязях, пластовых водах, сопровождающих нефтяные месторождения. В подзолах мало сульфатов, и биогенным путем сульфиды в них не накапливаются. В щелочных и нейтральных почвах образуются нерастворимые сульфиды. Накопление сульфида железа приводит к образованию черного ила.
С жизнедеятельностью сульфатвосстанавливающих бактерий связывают процесс биогенного содонакопления. Провинции, где формируются содовые солончаки, тяготеют к низменным местам, например, к поймам рек, понижениям вокруг озер. Если коренные породы в этих местах обогащены сульфатами, то в анаэробных условиях при затоплении и разложении органического вещества образуется H2S, который затем путем химических реакций преобразуется в Na2S и далее в соду при наличии карбонатов (СаС03, MgC03).
Биогенное появление соды не всегда сопровождается ее накоплением.
На гипсоносных отложениях не происходит содонакопления, так как гипс реагирует с содой и образуется вторичный сульфат натрия по схеме: Na2C03 + CaS04 Na2S04 + CaC03.
В присутствии тионовых бактерий, окисляющих Na2S, сода тоже не будет накапливаться. Таким образом, количественная сторона процесса содонакопления зависит от степени и длительности увлажнения почвы застойными водами, от количества органического вещества, характера отложений и наличия серо- окисляющих бактерий.
Восстановительные процессы в цикле серы не ограничиваются ассимиляционной и диссимиляционной сульфатредукцией. Восстанавливаться могут тиосульфаты и молекулярная сера. Восстановление S032~ и S2032" до S° проводят облигатно-анаэробные термофильные бактерии Thermoanaerobacterium thermosulfurigenes Clostridium thermosulfurogenes. Эти хемоорганотрофные бактерии, выделенные из термального источника в Йеллоустонском парке (США), вызывают брожение с образованием этанола, молочной и уксусной кислот и Н2; способны к гидролизу пектина и крахмала. Образующаяся при восстановлении тиосульфата молекулярная сера откладывается на клеточных стенках и выделяется в среду.
В восстановлении молекулярной серы до H2S участвуют многие термоацидофильные строго анаэробные архебактерии, например Thermoproteus tenax, Pyrococcus furiosus, Desulfurococcus mucosus и др. Эти бактерии — обитатели очень кислых сред с высокой температурой. Для Pyrococcus furiosus оптимальны: рН 1,0, температурный оптимум 100°С. В анаэробных условиях серу восстанавливают архебактерии Sulfolobus, которые в аэробных условиях входят в группу окислителей серы. И, наконец, бесцветные серные хемоорганогетеротрофные бактерии Beggiatoa alba в анаэробных условиях используют серу и тиосульфат в диссимиляцион- ной редукции как акцепторы электронов по типу анаэробного дыхания, сопряженного с окислением органических субстратов.
Сероводород образуется в больших количествах при разложении белковых веществ. Этот процесс, сопровождающийся выделением других летучих дурно пахнущих продуктов, обычно называют гниением. Гнилостные микроорганизмы могут быть аэробными и строго анаэробными. В основном это обычные сапротрофы родов Bacillus, Clostridium и др. Биогенная сера, поступающая в атмосферу в виде органических летучих соединений, в основном продукт деятельности гнилостных бактерий.
Процессы восстановительных звеньев цикла серы тесно сопряжены с окислительными, и часто сульфатредуцирующие бактерии развиваются в общих местообитаниях с серными, которые используют для окисления сероводород, поступающий из анаэробной зоны. Накопление в среде сероводорода может вызывать негативные последствия, так как он бывает причиной токсикоза почв, а в водоемах повышение концентрации H2S приводит к массовой гибели рыбы и других животных. В местах высокой активности сульфатвосстанавливающих бактерий происходит коррозия металлических конструкций в почве (H2S окисляет железо).
Помимо биологического цикла серы в атмосфере Земли происходят превращения серосодержащих газов без участия живых организмов. Сера в виде сернистого газа образуется в результате сжигания угля и нефти, а также при извержении вулканов в форме H2S. Эти соединения окисляются в атмосфере, а образующаяся серная кислота возвращается на поверхность земли и в почву (кислотные дожди). Вокруг промышленных районов таким путем на сотни и тысячи километров происходит повреждение растительности, особенно лесов, подкисление почв, иногда полная гибель растительности и образование так называемых «техногенных пустынь» вблизи химических предприятий.
|













|
К содержанию книги: Почвоведение - биология почвы
|
Круговорот серы в природе
Последние добавления:
Происхождение и эволюция растений
Биографии ботаников, биологов, медиков
Книги по русской истории Император Пётр Первый