|
|
Почвенные организмы |
Ассоциативная азотфиксация в биогеоценозах
|
Смотрите также:
Мейен - Из истории растительных династий
Биографии биологов, почвоведов
|
Московский государственный университет им. М. В. Ломоносова
Методы определения азотфиксации
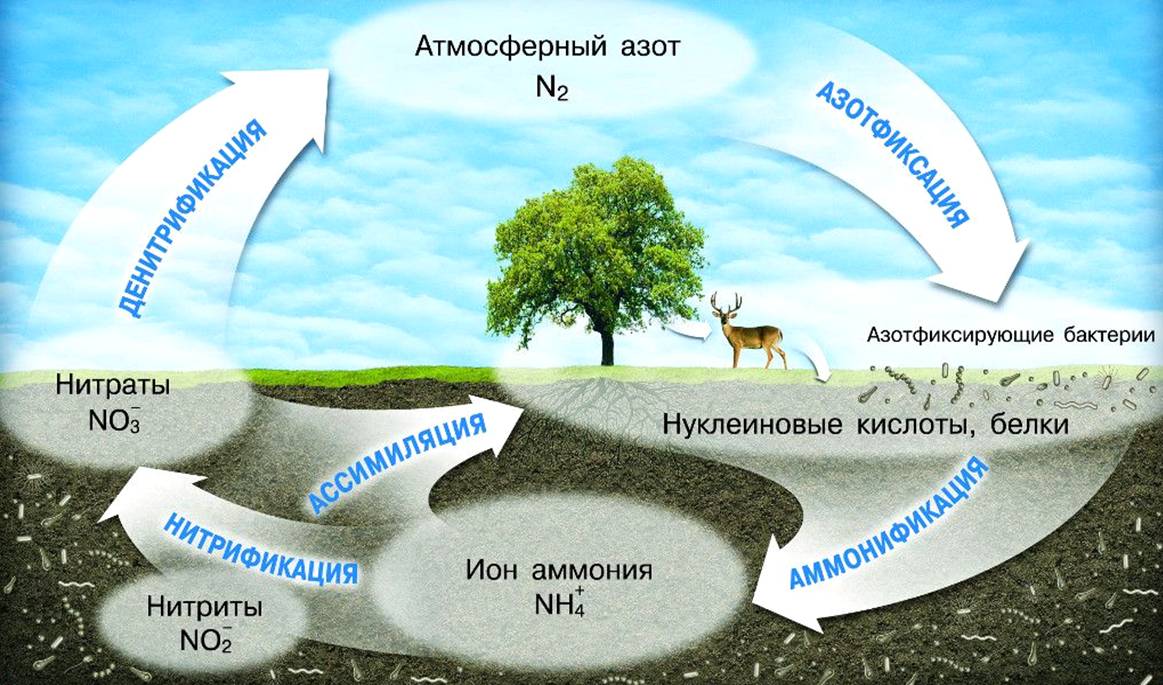
В последние годы наряду с большими успехами в области биохимии, физиологии и генетики азотфиксации существенно расширились и знания по экологии этого процесса.
Наиболее важным достижением явилось обнаружение повышенной азотфиксирующей активности в фитоплане (ризосфере и филлосфере) небобовых растений. Об активизации многих микробиологических процессов в прикорневой зоне растений известно давно. С появлением новых методов определения азотфиксации было установлено, что и азотфиксирующая активность микроорганизмов существенно выше вблизи и на поверхности растений, чем в почве без растений.
К настоящему времени накоплен обширный экспериментальный материал, убедительно свидетельствующий о широком распространении фитоплановой азотфиксации у растений различных семейств и в разных биогеоценозах. В отличие от известных форм азотфиксации ее принято называть ассоциативной азотфиксацией.
Несмотря на тс, что при ассоциативной азотфиксации микроорганизмы и растения не вступают в столь тесное взаимодействие, которое характерно для симбиотических систем, в целом она характеризуется примерно такими же экологическими особенностями. Было выявлено, в частности, что ассоциативная азотфиксация имеет хорошо выраженную сезонную динамику, связанную с развитием растений в течение вегетации. Она нарастает с появлением проростков и по мере их роста, достигает максимума в период цветения, постепенно снижается ко времени созревания и резко падает после уборки урожая или естественного отмирания растительности (Ума- ров, 1982; Boddey, Dobereiner, 1982). Другим свойством ассоциативной азотфиксации является ее суточная периодичность, зависящая от циркадных ритмов растений (Balandreau et al., 1976; Watanabe et al., 1980). В отсутствие растений (черный пар, междурядья в посевах, кальвиции и др.) таких изменений азотфиксирующей активности в почве не наблюдается. '
Весьма важным для понимания структуры и продуктивности биогеоценозов было выяснение того, что суммарный уровень, продуктивность азотфиксации при налчии растений значительно выше, нежели в их отсутствие. Согласно оценочным данным, масштабы ассоциативной азотфиксации в биогеоценозах зоны умеренного климата составляют в среднем 25—55 кг/га за вегетационный период (Мишустин и др., 1978), а в максимуме могут достигать 50—150 кг/ га (Умаров, 1982).
Оценка продуктивности азотфиксации
Точная оценка продуктивности ассоциативной азотфиксации в конкретных биогеоценозах может быть проведена на основе многократных прямых измерений активности процесса в течение периода вегетации и при учете его пространственной неоднородности. Ниже приведены некоторые из результатов, полученных таким способом в агробиогеоценозах на дерново-подзолистых почвах (кг/га, среднее за вегетационный период 1979 г.). Под посевами ячменя и луговых злаков суммарная продуктивность азотфиксации за вегетационный период составила около 40 кг/ia, под картофелем — около 30 кг/га. В то же время в почве под паром и в междурядьях она была на уровне 10—13 кг/га.
Различия в продуктивности азотфиксации на участках, занятых растениями, и на парующей почве позволяют выявить соотношение между ассоциативной азотфиксацией, протекающей за счет энергии корневых выделений и корневого опада, и несимбиотнческой азот- фиксацией, осуществляющейся главным образом благодаря энергии корневых и пожнивных растительных остатков. В изучавшихся агробиогеоценозах на долю несимбиотнческой азотфиксации приходится около 25—30% от суммарной продукции азотфиксации. Можно предполагать, что указанное соотношение будет сохраняться и в других биогеоценозах; вероятно, ассоциативной азотфиксации принадлежит значительная роль в азотном балансе почв.
Другая отличительная особенность ассоциативной азотфиксации — отсутствие резких колебаний в численности микроорганизмов, проводящих этот процесс. Она протекает при относительно постоянном количестве микрооргаиизмов-диазотрофов в ризосфере (Мошко- ва, 1980). Вклад азотфиксирующих бактерий в азотный баланс почв при ассоциативной азотфиксации связан не с общей их биомассой в фитоплане, а с их активным физиологическим состоянием в периоды быстрого развития растений. Прикорневая зона в такие периоды может рассматриваться в виде своеобразного проточного культиватора микроорганизмов, обеспечивающего эффективное протекание микробиологических процессов.
Как уже отмечалось, ассоциативная азотфиксация осуществляется и в филлосфере. Она была обнаружена в филлосфере большого числа растений (табл. 1), причем активность ее колебалась от 0,12 до 7,01 мг азота Ю-1 м2/ч~* (Садыков, 1980).
Изучение сезонной динамики ассоциативной азотфиксации на листьях и стеблях березы и тимофеевки показало, что суммарная продуктивность ее в филлосфере этих растений достигает 7—10 кг/га (Садыков, Умаров, 1980а).
Путем одновременного раздельного определения величин азотфиксации в ризосфере и филлосфере одних и тех же видов растений удалось выяснить, что уровень ассоциативной азотфиксации при расчете на единицу поверхности примерно одинаков (табл. 2).
Таким образом, растения являются главным фактором, определяющим динамику и продуктивность ассоциативной азотфиксации в природе.
Основным энергетическим субстратом для азотфиксирующих бактерий в фитоплане растений служат продукты экзоосмоса и корневой опад. По последним оценкам, суммарная величина этих продуктов, представленных в основной массе полисахаридами, составляет за вегетационный период около 7з от общей продукции фотосинтеза (Sauerbeck et al., 1982). Массированное поступление полисахаридов в фитоплан приводит к усилению ассоциативной азотфиксации. Некоторые факты позволяют полагать, что по крайней мере одна треть этих соединений прямо расходуется на ассоциативную азотфиксацию.
При активной азотфиксации микроорганизмы могут выделять разнообразные азотсодержащие метаболиты, преобладающими среди которых являются аминокислоты. Помимо прижизненного выделения аминокислот, азотфиксаторы могут обогащать ими ризосферу и филлосферу и постмортально, после гибели клеток и аммонификации белков. Вероятно, этот процесс имеет подчиненное значение, поскольку, как отмечалось, численность микроорганизмов в фитоплане остается сравнительно постоянной в течение всего периода вегетации.
Таким образом, чем более развитым фотосинтетическим аппаратом обладает растение и, следовательно, чем больше энергии фотосинтеза транспортируется им в ризосферу и филлосферу, тем выше уровень ассоциативной азотфиксации и больше азотсодержащих соединений поступает в фитоплан. Приток азота в растение при этом находится в соответствии с интенсивностью образования в нем реакционно-способного углеродного материала, что является одним из главных условий сбалансированного роста.
Зависимость ассоциативной азотфиксации от активности фотосинтетической деятельности растения
Зависимость ассоциативной азотфиксации от активности фотосинтетической деятельности растения свидетельствует о тесном сопряжении двух уникальных биологических процессов — азотфиксации и фотосинтеза — не только у симбиотически фиксирующий систем, но и у широчайшего круга растений, обеспечивая им определенную автономность в азотном питании. Можно полагать, что именно в этом одна из причин устойчивости и высокой продуктивности климаксных фитоценозов, не имеющих обычно в своем соста* ве бобовых растений.
В отношении азотфиксирующих бактерий, ассоциированных с растениями, пока нет единого мнения. Неоднократно появлялись сообщения о приуроченности определенных видов бактерий к ризосфере различных травянистых растений (Boddey, Dobereiner, 1982), Отмечалось, что в ризосфере сорго, риса, маиса, пшеницы и ряда тропических пастбищных злаков широко распространены бактерии рода Azospirillum (Dobereiner, 1974), а в ризосфере сахарного тростника — бактерии род% Beijerinckia. У зерновых злаков нередко обнаруживаются представители энтеробактерий и бацилл (Larson, Neal, 1976).
Были высказаны предположения о том, что такие ассоциаций между определенными видами бактерий и растений достаточно тесны, чтобы считать их высокоснецифичными. По миению И. Доберей- нер (Dobereiner, 1978), существуют малоспецифичные ассоциации так называемого экторизосферного типа и высокоспецифичные ассоциации эндоризосферного типа, различающиеся по силе адсорбции бактерий на поверхности корня. Если первые сравнительно легко удаляются даже водой, то при втором типе взаимодействия микроорганизмы локализованы в слое муцигеля и их не удается смыть даже путем применения поверхностно-активных веществ. Более того, имеются данные (Dazzo, Hubbell, 1974) о наличии бактерий такого типа и в тканях корня.
Выводы Доберейнер и других исследователей о высокой специфичности ассоциаций диазотрофных бактерий с корнями определенных растений оспариваются (Brown, 1976; Tyler et al., 1979). Показано, что разные азотфиксирующие бактерии способны образовывать тесные ассоциации с корнями одних и тех же растений (Staphorst, Strijdom, 1978), а специфическая ассоциация с корнями растений не является обязательным условием высокой активности азотфиксации (Tjepkema, Burris, 1976).
По нашим наблюдениям, в ризосфере различных растений доминируют представители родов Bacillus, Azotobacter, Clostridium, Beijerinckia, Arthrobacter, Spirillum и коринеподобные бактерии. Наиболее часто встречаются Вас. polymyxa, Вас. circulans, Pseudo- monas denitrificans и коринеподобные бактерии. Не отмечено какой- либо специфичности ни в отношении определенных видов бактерий, ни в отношении видов растений.
Пока мало сведений об азотфиксирующих микроорганизмах в филлосфере, а имеющиеся данные свидетельствуют об отсутствии специфических видов на поверхности листьев и стеблей растений (Jones, 1976; Meyer, Todd, 1976).
Для филлосферы всех изученных нами растений было характерно преобладание пигменто- и слизеобразующих видов бактерий и дрожжей (Садыков, Умаров, 1980). При посеве из смывов листьев и при анализе методом отпечатков не удалось выявить различий в составе бактерий в филлосфере березы и тимофеевки. В течение всего периода вегетации доминировали представители родов Erwinia, Pseudomonas, Corynebacterium и Arthrobacter. Редко обнаруживались спороносные бациллы и дрожжи.
Анализ имеющихся к настоящему времени данных позволяет заключить, что ассоциативная азотфиксация осуществляется смешанной популяцией различных микроорганизмов-гетеротрофов, обладающих высокой азотфиксирующей активностью в условиях проточного культивирования. Способностью к азотфиксации обладают большинство, если не все, почвенные бактерии (Умаров, 1982; Uma- rov, Zvyagintsev, 1982). Правомочность этого вывода основывается также и на том, что ни один из важнейших процессов природных процессов не осуществляется узкой группой микроорганизмов. Азот- фиксация наблюдается в почвах всех географических зон, протекает в илах, донных осадках, водоемах, в ризосфере и филлосфере всех растений, в кишечнике насекомых, рыб и высших животных, т. е. ЕО всем многообразии экосистем в природе. Устаревшим следует считать взгляд на азотфиксацию как уникальный процесс, осуществляющийся лишь узкой группой высокоспециализированных организмов (Умаров, 1982). Действительно, список азотфиксирующих бактерий непрерывно растет (Postgate, 1978). В него включены не только клубеньковые бактерии вне симбиоза с бобовыми растениями (Мишустин, Громыко, 1975), но и почвенные актиномицеты, у которых способность к азотфиксации не удавалось установить в течение длительного времени (Gauthier et al., 1981).
Способность к фиксации молекулярного азота у микроорганизмов
В последнее время появились данные, прямо свидетельствующие о широком распространении способности к фиксации молекулярного азота у микроорганизмов. Так, около 71 % изолятов бактерий, выделенных из отвалов рекультивированных шахт, обладали способностью редуцировать ацетилен (Коропеп et al., 1980). Нитрогеназ- ная активность была обнаружена более чем у 55% штаммов филло- сферных бактерий (Садыков, 1980).
В табл. представлены данные о соотношении азотфиксирующих бактерий в общем числе микроорганизмов, вырастающих на безазотных питательных средах. Свыше 60% изолятов показали наличие азотфиксирующей активности. Одновременно было установлено, что доля азотфиксирующих бактерий примерно вдвое выше в ризосфер- ной почве по сравнению с почвой, отобранной вдали от корней. Преобладание бактерий-азотфиксаторов в ризосфере косвенно подтверждает их важную роль в азотном питании растений.
В этой связи уместно отметить неоднократные предложения генетиков решить проблему снабжения растений «биологическим» азотом путем передачи гена азотфиксации от Klebsiella pneumoniae бактериям из ризосферы важнейших сельскохозяйственных небобо-] эвых растений (Клингмюллер и др., 1981). Из-за отсутствия видовой
Соотношение азотфиксирующих и нефйксирующих микроорганизмов в некоторых почвах Кавказа в течение вегетационного периода специфичности между бактериями и растениями, а также вследствие природного многообразия азотфиксирующих микроорганизмов следует признать бесперспективность таких подходов к решению указанной проблемы.
Представляются спорными и предложения осуществить искусственную бактеризацию небобовых растений высокоактивными штаммами азоспириллы (Dobereiner, 1978) или слабовирулентными культурами фитопатогенных бактерий. Как известно, такие попытки неоднократно предпринимались в прошлом (азотобактерин), но не дали положительных результатов с точки зрения обогащения почвы и растений дополнительным количеством азота; даже при активном размножении азотобактера в почве его благоприятное действие на развитие растений было связано с продуцированием им биологически активных веществ (Мишустин, 1979).
Более перспективным представляется возможность активизации ассоциативной азотфиксации путем усиления фотосинтетической деятельности растений и за счет оптимизации экологических факторов.
Среди многообразных факторов окружающей среды, влияющих как на активность фотосинтеза, так и на азотфиксирующую активность микроорганизмов, существенное значение имеет влажность почвы и атмосферы. Как известно, влажность почвы, выражающая содержание воды в процентах от массы сухой почвы, лишь косвенно характеризует физиологическую доступность ее для живых организмов. При одной и той же влажности вода, содержащаяся в песчаной почве, легко доступна растениям и микроорганизмам, тогда как в глинистой почве — совершенно недоступна, что обусловлено различной величиной и качеством поверхности почвенных частиц. Более универсальными, прямо определяющими физиологическую доступность влаги в почве являются термодинамические характеристики — потенциал или давление почвенной влаги (Судницын, 1979).
К сожалению, несмотря на обилие в литературе данных о влиянии различной влажности почвы на активность азотфиксации, получены они без учета фактической доступности влаги для микроорганизмов. Отсутствуют данные об оптимальных значениях влажности для ассоциативной азотфиксации. Зависимость активности ассоциативной азотфиксации от давления почвенной влаги была исследована для дерново-подзолистой тяжелосуглинистой почвы (Мошкова и др., 1982).
Так как в природных условиях детально изучить зависимость активности азотфиксации от влажности почвы невозможно из-за одновременного действия сложного комплекса факторов, в модельном эксперименте давление влаги изменяли в пределах от —3,65 до —0,09 МПа.
Полученные данные показали четко выраженное стимулирующее влияние повышенного содержания влаги в почве на уровень азотфик- сирующей активности. В целом зависимость азотфиксации от давления почвенной влаги представляет собой S-образную кривую. При увеличении влажности от 8 до 19% (от —3,65 до —1,41 МПа) активность азотфиксации изменяется незначительно — от 0,72 до 0,90 мг азота/кг почвы «ч"1. При дальнейшем увеличении содержания влаги в почве активность резко возрастает — в интервале от —1,3 до —0,24 МПа примерно в 50 раз. Затем рост активности замедляется.
Закономерности, установленные при изучении азотфиксации в почве в модельных условиях, были подтверждены и в вегетационных опытах с растениями овсяницы и тимофеевки. При проведении этих экспериментов температура почвы и освещенность растений были постоянны, продолжительность каждой серии опытов невелика (не более недели). Во всех вариантах использовали хорошо развитые, активно веге- тирующие растения. Поэтому изменения уровня азотфиксации определялись главным образом содержанием влаги в почве.
Зависимость активности ассоциативной азотфиксации от давления почвенной влаги в условиях вегетационного опыта оказалась аналогичной зависимости, полученной в модельных опытах ( 1). Максимум активности соответствовал давлению влаги —0,25 МПа, а начало резкого подъема ее наблюдалось при — 1,4 МПа ( 1).
Интересно отметить, что последняя величина близка к значению потенциала влаги в почве, при котором начинает возрастать и транс- пирация у растений (Судницын, 1979). Не исключено, что эта величина отражает достаточно общие свойства живых организмов, как растений, так й почвенных микроорганизмов, по отношению к воде в окружающей среде. В полевых условиях для выяснения степени влияния влажности почвы на ассоциативную азотфиксацию сопоставляли значения активности азотфиксации со значениями влажности почвы.
Хотя в природных (полевых) условиях на действие фактора влажности накладывается действие сложного комплекса многих других факторов, определяющих уровень фотосинтеза и азотфиксации (погодные условия, освещенность, содержание соединений азота в почве, концентрация С02 в воздухе и др.), тем не менее даже на этом динамичном в течение вегетационного периода фоне хорошо прослеживается стимулирующее влияние повышенной увлажненности почвы на уровень ассоциативной азотфиксадии. Корреляционный анализ результатов позволил установить, что существует тесная связь между влажностью почвы и значениями азотфиксирующей активности (табл. 4).
Таким образом, результаты лабораторных, вегетационных и полевых опытов убедительно свидетельствуют о высокой степени влияний влажности почвы на ассоциативную азотфиксацию.
Азотфиксирующая активность в филлосфере
Оптимизация влажности почвы может позволить регулировать активность ассоциативной азотфиксации в агробиогеоценозах — поддерживая увлажненность почвы на необходимом уровне, можно повысить обеспеченность растений «биологическим» азотом. Изучение азотфиксации в дерново-подзолистых почвах при различных температурных режимах показало, что температура почвы не играет заметной роли в активизации ассоциативной азотфиксации (табл. 4). При сопоставлении значений ассоциативной азотфиксации в ряде агробиогеоценозов с изменениямй температуры верхних горизонтов (О—10 см) почвы корреляции не, установлено. Обусловлено это тем, что в течение летних месяцев температура почвы колеблется не столь значительно, как влажность. Кроме того, лимитирующим фактором для фотосинтеза и азотфиксации температура почвы становится лишь при относительна низких значениях — ниже 7° (Чундерова и др. 1974; Jlapxep, 1978). Лётом температура почвы в верхних горизонтах в средней полосе редко опускается ниже 10° (по многолетним наблюдениям Метеорологической обсерватории МГУ средняя температура почвы за 3 летних месяца колебалась в пределах от 12 до 19°).
На азотфиксирующую активность в филлосфере оказывают влияние температура и влажность воздуха, а также освещенность растений. Температура начинает достоверно снижать активность азотфиксации при значениях ниже 20°, относительная влажность воздуха — ниже 60%, а освещенность — менее 25 клк. Азотфиксация в филлосфере полностью прекращается при температуре воздуха около 2°, относительной влажности воздуха менее 15%. Предложено соотношение, позволяющее производить расчет активности ассоциативной азотфиксации (FaKT) в филлосфере по известным величинам потенциальной активности азотфиксации (FnoT) с учетом факторов температуры (Ft), влажности (Fw) воздуха ц освещенности растений (Fe) (Садыков, Умаров, 19806)
Сопоставление данных, полученных непосредственно в природе путем прямых определений, с результатами расчетов показало хорошее соответствие, что свидетельствует о том, что в формуле учтены наиболее существенные факторы внешней среды, влияющие на уровень ассоциативной азотфиксации в филлосфере.
Применимость показателей г гw и Fe, полученных для филло- сферы тимофеевки, для расчета активности азотфиксации в филлосфере березы позволяет предполагать, что эти зависимости носят универсальный характер (Садыков, Умаров, 19806)*.
Азотфиксирующая активность в ризосфере и филлосфере существенно зависит от концентрации С02 в атмосфере над растением. Как было показано в последние годы (Ковда, 1981), продуктивность фи- тоценозов не в последнюю очередь определяется содержанием С02 в воздухе. Считается, что именно нехватка углекислоты для нормального протекания фотосинтеза — одна из причин снижения продуктивности агробиогеоценозов при длительном одностороннем применении минеральных удобрений (Ковда, 1981).
Согласно расчетам И. С. Шатилова (1981), в период интенсивного развития растения синтезируют около 360—380 кг/га сухой массы за сутки, затрачивая до 720 кг С02. В то же время в слое воздуха над гектарным участком содержится около 5 кг С02; необходим постоянный источник углекислоты и интенсивный газообмен для поддержания высокой продуктивности фотосинтеза. По данным дабораторных и полевых наблюдений, интенсивность и продуктивность фотосинтеза заметно увеличиваются с возрастанием концентрации С02 в воздухе от 300—320 до 500—700 частей на 1 млн. (Ковда, 1981).
Повышение концентрации углекислоты в атмосфере должно приводить не только к усилению фотосинтетической деятельности растений, но и к активизации ассоциативной азотфиксации. Действительно, как было показано в модельных опытах с «гнотобиотическими системами», состоявшими из стерильных проростков риса, инокулиро- ванных смешанными азотфиксирующими культурами микроорганизмов, введение в атмосферу инкубационной камеры дополнительного количества С02 вызывало увеличение нитрогеназной активности в системе ( 2).
Активное регулирование концентрации углекислоты в атмосфере (внесение навоза, компостов. запашка соломы и растительных остатков) благоприятно не только для процессов фотосинтеза, но и способствует возрастанию доли «биологического» азота в урожае.
Влияние минеральных соединений азота на процесс фотосинтеза у растений и азотфиксацию у микроорганизмов
Одним из факторов среды, оказывающих сильное регулирующее действие на ассоциативную азотфиксацию, являются минеральные азотные удобрения. В литературе имеются противоречивые сведения о влиянии минеральных соединений азота на активность азотфиксации в почве. Широко распространено мнение, что минеральный азот сильно тормозит этот процесс. В то же время имеются данные (Калининская и др., 1977; Мишустин и др., 1977; Watanabe et al., 1981), что дозы минерального азота, обычно используемые в сельскохозяйственной практике (100—150 кг на 1 га), вызывают лишь кратковременное подавление азотфиксации, а существенное торможение наблюдается лишь при дозах связанного азота в 500—1000 кг/га. Характерно также, что азотные удобрения действуют в первую очередь на фото- синтезирующие бактерии-азотфиксаторы и только с увеличением дозы ингибируют азотфиксирующую активность гетеротрофных бактерий.
Влияние минеральных соединений азота на процесс фотосинтеза у растений и азотфиксацию у микроорганизмов прямо противоположно. Фотосинтез протекает наиболее активно, если имеется сбалансированность между поступлением углерода и азота в растительный организм. При наличии в прикорневой зоне достаточного количества легкодоступных соединений азота растения используют их в качестве главного источника азотного питания. Однако концентрация минеральных соединений азота в почве не остается постоянной — помимо .потребления растениями, они иммобилизуются в телах почвенных микроорганизмов, выносятся в грунтовые воды, закрепляются в почве и, что следует особо подчеркнуть, подвергаются денитрификации.
К настоящему времени можно считать твердо доказанным, что скорость и масштабы денитрификации значительно выше в ризосфере растений (Куракова, Умаров, 1982, Scott, Tiedje, 1979), а основные дотери азота из почвы происходят в газообразной форме (Смирнов и др., 1981; Watanabe et al., 1981). В результате совокупного действия всех указанных факторов почва быстро обедняется доступным азотом, причем наиболее сильно — в прикорневой зоне, так как интенсивность микробиологических процессов трансформации азота максимальна в ризосфере (Илялетдинов, 1976). Закономерно также, что скорость протекания этих процессов определяется в целом уровнем бдологической активности или окультуренности почв. В почвах с высокой биологической активностью потери азота выше и, как следствие, ниже коэффициент использования азотных удобрений; в таких почвах быстрее наступает состояние азотного дефицита (Сапожников и др., 1978). Таким образом, чем выше биологическая активность почв и чем лучше развиваются растения, тем быстрее будет исчерпан запас легкодоступных соединений азота в прикорневой зоне, тем быстрее растения окажутся в условиях глубокого азотного голодания. При нарушении сбалансированности между поступлением углерода и азота в растительный организм возрастает эксуда- ция как ответная реакция на ухудшение условий среды (Сытиик и др., 1972). Снижение концентрации минеральных соединений азота в прикорневой зоне и увеличение масштабов экзоосмоса способствуют усилению ассоциативной азотфиксации. При этом чем в большей дозе вносились в почву азотные удобрения и соответственно лучше развивались растения, тем выше активность и суммарная продуктивность ассоциативной азотфиксации.
Вероятно, именно в усилении азотного питания за счет активизации ассоциативной азотфиксации заключается экологический смысл массированной эксудации растениями. Этот своеобразный экологический саморегулирующийся механизм связывает высшие растения и ассоциированные с ними микроорганизмы, в том числе и азотфик- сирующие.
Азотные удобрения служат в качестве «пускателя;) такого механизма; их можно рассматривать в виде «стартового азота» роль которого хорошо известна для азотфиксирующих микроорганизмов. Усиление ассоциативной азотфиксации в фитоплане растений способствует снижению коэффициента использования азотных удобрений. Именно этим можно объяснить хорошо известное в агрохимии явление снижения коэффициента использования минеральных азотных удобрений по мере увеличения вносимых в почву доз (Петербургский, 1979). Следовательно, оптимизация доз азотных удобрений с учетом свойств почвы, биологических особенностей растения и экологии ассоциативной азотфиксации может позволить увеличить долю «биологического» азота в урожае и более экономно расходовать минеральные удобрения. Лишь малой изученностью процессов взаимодействия растений и азотфиксирующих микроорганизмов можно объяснить отсутствие практических рекомендаций с целью достижения оптимального соответствия между азотом «биологическим» и минеральным в продукции растениеводства. Раскрытие тонких особенностей такого взаимодействия позволит также понять функциональную роль каждого из компонентов биогеоценоза.
Выводы
Растения являются ведущим экологическим фактором, определяющим динамику и продуктивность ассоциативной азотфиксации в биогеоценозах. Ассоциативная азотфиксация осуществляется смешанной популяцией азотфиксирующих бактерий. Способностью к азотфиксации обладает большинство почвенных бактерий.
Ассоциативная азотфиксация наиболее активно протекает при давлении почвенной влаги около — 0,25 МПа. Температура почвы не оказывает заметного влияния на динамику азотфиксации.
Регулирование активности ассоциативной азотфиксации возможно путем оптимизации доз минеральных азотных удобрений, концентрации углекислоты в атмосфере.
ЛИТЕРАТУРА
Илялетдинов А. Я. Микробиологические превращения азотсодержащих соединений в почве. Алма-Ата: Наука, 1976. 281 с. Калининская Т. А., Миллер Ю. МРао В. Р., Белов Ю. М. Активность несим- биотической азотфиксации в почвах рисовых полей.— В кн.: Повышение плодородия почв рисовых полей. М.: Наука, 1977, с. 97—105. Клингмюллер ВБахейтел Д., Хофманн А. Генетическое манипулирование с признаком азотфиксации у почвенных бактерий.— В кн.: Молекулярные основы генетических процессов. М.: Наука, 1981, с. 239—247. Ковда В. А. Советское почвоведение па службе сельского хозяйства СССР. Тбилиси, 1981. 106 с. Куракова Я. Г., Умаров М. М. Влияние растений на сопряженное протекание процессов азотфиксации и денитрификации.— Вести. МГУ. Сер. 17. Почвоведение, 1982, № 3, с. 67—69. Лархер В. Экология растений. М.: Мир, 1978. 185 с. Мишустин Е. Н. Газообразные соединения азота в почве.— В кн.: Роль микроорганизмов в круговороте газов в природе. М.: Наука, 1979, с. 68—76. Мишустин Е. Я., Громыко Е. П. Азотфиксация у клубеньковых бактерий вне симбиоза.— Изв. АН СССР. Сер. биол., 1975, № 5, с. 637—640. Мишустин Е. Я., Калининская Т. АПетрова А. Я. Влияние альголизации на урожай риса.— В кн.: Повышение плодородия почв рисовых полей. М.: Наука, 1977, с. 204—212. Мишустин Е. Н., Калининская Т. АЧерепков Я. И. Несимбиотическая азотфиксация в пахотных почвах.— В кн.: Проблемы почвоведения. М.: Наука, 1978, с. 92—96. Мошкова М. В. Несимбиотическая азотфиксация в агроценозах на дерново-подзолистой почве: Автореф. дис. ... канд. биол. наук. М.: МГУ, 1980. 25 с. Мошкдва М. В., Судницын И. Ж, Умаров М. М. Влияние влажности на азотфик- сирующую активность дерново-подзолистых почв.— Почвоведение, 1982, № 1, с. 92-95. Петербургский А. В. Круговорот и баланс питательных веществ Е земледелии М.: Наука, 1979. 47 с. Садыков Б. Ф. Азотфиксация в филлосфере растений: Автореф. дис. ... канд биол. наук. М.: МГУ, 1980. 23 с. Садыков Б. Ф., Умаров М. М. Обнаружение азотфиксирующей активности в филлосфере растений.— Микробиология, 1980а, т. 49, вып. 1, с. 147—150. Садыков Б. Ф., Умаров М. М. Продуктивность азотфиксации в филлосфере растений.— Микробиология, 19806, т. 49, вып. 6, с. 998—1002. Сапожников Н. А., Ливанова Т. КРусинова И. ПСирота Л. Б. Трансформация азота в дерново-подзолистых почвах различной окультуренности и поступление азота в растения.— В кн.: Проблемы почвоведения. М.: Наука, 1978,. с. 114—120. Смирнов П. М., Кидин В. ВИванникова Л. А. Превращение продуктов нитрификации в почве и их роль в газообразных потерях азота удобрений.— Изв. АН СССР. Сер. биол., 1981, № 3, с. 370—383. Судницын И. И. Движение почвенной влаги и водопотребление растений. M.i Изд-во МГУ, 1979. 253 с. Сьгтник К. М., Книга Н. ММусатенко Л. И. Физиология корня. Киев: Наук, думка, 1972. 225 с. Умаров М. М. Значение несимбиотической азотфиксации в балансе азота в почве.— Изв. АН СССР. Сер. биол., 1982, № 1, с. 92—105. Умаров М. М., Куракова Н. Г. Сопряженное изучение азотфиксации и денитрификации в почве.— Вестн. МГУ. Сер. 17. Почвоведение, 1982, № 3, с. 56—58. Чундерова А. ИЗубко И. К., Князева В. Л. Влияние физико-химических факторов среды на численность и активность азотфиксирующих микроорганизмов.— В кн.: Динамика микробиологических процессов в почве. Таллин: Изд-во АН ЭССР, 1974, ч. II, с. 14—17. Balandreau Rinaudo G.y Umarov M., Dommergues Y. A non-symbiotic nitrogen fixation in paddy soils.— In: Proc. 1st Intern, symp. on N2 fixation. Wash.: Univ. press, 1976, p. 611—628. Boddey R., Dobereiner J. Association of Azospirillum and other diazotrophs with tropical Gramineae.— In: Non-symbiotic nitrogen fixation and organic matter in the tropics. Symposia papers 1. New Delhi, 1982, p. 28—47. Brown M. Role of Azotobacter paspalii in association with Paspalum notatum.— J. Appl. Bacteriol., 1976, vol. 40, N 3, p. 341—348. Dazzo F., Hubbell D. Biological nitrogen fixation.— Soil and Crop Soc. Florida proc., 1974, vol. 34, p. 71—79. Dobereiner J. Nitrogen-fixing bacteria in the rhizosphere.— In: Biological nitrogen fixation. Amsterdam etc., 1974, vol. 33, p. 86—120. Dobereiner J. Nitrogen fixation in grass-bacteria associations in the tropics.— In: Isotopes Biol, dinitrogen fixat. proc. Vienna, 1978, p. 51—68. Gauthier D.y Diem H., Dommergues Y. In vitro nitrogen fixation by two actino- mycete strains, isolated from casuarine nodules.— Appl. Environ. Microbiol., 1981, vol. 40, N 1, p. 300-308. Jones K. Nitrogen fixing bacteria in the canopy of conifers in a temperate forest.— In: Microbiology of aerial plant surfaces. London; New York: Acad, press, 1976, p. 73-81. Koponen M., Funke В., Galitz D. Enumeration of acetylene-reducing bacteria in strip-mined reclamation sites in a temperate grassland.— Plant and Soil, 1980, vol. 57, N 2/3, p. 399—403. Larson R., Neal J. Selective colonization of the rhizosphere of wheat by nitrogen-fixing bacteria.— Ecol. Bull., 1976, vol. 26, p. 331—341. Meyer R., Todd R. Epiphytic nitrogen fixation in a forest ecosystem.— In: Abstr. ann. meet. Amer. Soc. microbiol. Wash., 1976, p. 171—179. Postgate J. Biological nitrogen fixation.— In: Co. microbiol. selec. Top. Further study. London; New York, 1978, p. 343—361. Sauerbeck D., Nonnen S., Allard J. Consumption and turnover of photosynthates in the rhizosphere.— Abstr. 12 Congr. ISSS, India, New Delhi, 1982, p. 59. Scott M., Tiedje J. The effect of roots on soil denitrification.— Soil sci. soc. Amer. J., 1979, vol. 43, N 5, p. 951—956. Staphorst J., Strijdom B. Diazotrophic bacteria associated with pasture and veld grasses in South Africa.— Phytophylactica, 1978, vol. 10, N 1, p. 13—16. Tjepkema J., Burris R. Nitrogenase activity associated with some Visconsin prairie grasses.— Plant and Soil, 1976, vol. 45, N 1, p. 81—94. Tyler M., Milam /., Smith R. et al. Isolation of Azospirillum from diverse geographic regions.—Canad. J. Microbiol., 1979, vol. 25, N 6, p. 693—697. Vmarov M., Zvyagintsev D. Role of non-symbiotic nitrogen fixation in soil nitrogen budget.— Abstr. 12 Congr. ISSS, India, New Delhi, 1982, p. 311. Watanabe J., App A., Alexander M. Non-symbiotic nitrogen fixation associated with the rice plant in flooded soils — Soil Sci., 1980, vol. 130, N 5, p. 283—289. Watanabe /., Guzman M., Cabrera D. The effect of nitrogen fertilizer on N^-fixa- tion in the paddy field measured by in situ acetylene reduction assay.— Plant and Soil, 1981, vol. 135, N 1, p. 135—139. |











Ассоциативная азотфиксация в биогеоценозах
К содержанию книги: Почвенные организмы как компоненты биогеоценоза