|
|
Почвенные организмы |
Актиномицеты в биогеоценозах
|
Смотрите также:
Мейен - Из истории растительных династий
Биографии биологов, почвоведов
|
Московский государственный университет им. М. И. Ломоносова
Основная экологическая функция актиномицетов — разложение органического вещества
Для актиномицетов основным местом обитания является почва, а основной экологической функцией — разложение органического вещества на поверхности почвы и в ее минеральных горизонтах (Waksman, 1959). Однако среди этой группы организмов отмечены также возбудители заболеваний человека и животных, немногочисленные патогены растений (Lechevalier II. A., Lechevalier М. Р., 1981), продуценты антибиотиков (Okami, 1973; Waksman, Lechevalier, 1962). Отдельные представители актиномицетов могут нарушать деятельность человека, разлагая резиновые прокладки канализационных труб, утилизируя авиационное топливо, образуя пахучие вещества, загрязняющие питьевую воду, и т. д. (Lechevalier, 1974).
Почва — биотоп, в котором сосредоточивается основной пул актиномицетов. Многочисленные обзоры последних лет посвящены представителям рода Streptomyces (Lacey, 1973; Kuster, 1976; Williams, 1978; Kuntzner, 1981), экологии стрептомнцетов посвящены работы Вильямса с сотрудниками (Davis, Williams, 1970; Khan, Williams, 1975).
В последнее десятилетие литература пополнилась сведениями о распространении в природе представителей других родов порядка Actinomycetales. Актиномицеты выявлены во всех известных почвах мира, однако их численность, роль в биоценозах, биохимическая активность изменяется в зависимости от эколого-географнческих условий.
Эколого-географические закономерности распространения актиномицетов изучены в основном для представителей родов Strcptomyces и Streptoverticillium.
По данным Е. Н. Мишустина (1972, 1981), почвы северных климатических зон значительно беднее южных но числу актиномидетов. Если в почвах полярной тундры актиномицеты составляют всего 0,4—3,4% от общего количества бактерий (Мишустин, Мирзоева, 1953), в торфяниках и торфяно-болотных почвах — 1,8—2,7% (Кра- сильников и др., 1953; Жданникова, 1963) и не являются доминирующими формами среди бактерий в подзолистых почвах (Рыбалки- на, Кононенко, 1957; Аристовская, 1965; Зенова, Вустина, 1979), то в черноземах, каштановых почвах, желтоземах влажных субтропиков, сероземах актиномицеты представлены разнообразными видами и составляют 20—40% от общего количества бактерий (Новогруд- ский, 1946; Чулаков, 1960; Егорова, 1966; Рыбалктша, Кононенко, 1957; Андреюк и др., 1974; Зенова, Вустина, 1980). Богаты актино- мицетами горные почвы, особенно почвы альпийских и субальпийских лугов (Андреюк и др., 1974); в песчаио-пустынных почвах актиномицеты по численности превосходит бактерии (Клевенская, 1960; Зенова, Вустина, 1980; Мишустин, 1975).
Стрептомицеты в разных почвах имеют либо различный набор групп и видов, либо сходный спектр, но с разным соотношением численности составляющих его популяций. Групповой спектр стреп- томицетов и разнообразие их видового состава значительно расширяется от почв северных зон к южным. Исключение составляют солончаки и красноземы. Стрептомицеты в солончаках представлены преимущественно группой, лишенной воздушного мицелия; красноземы при общей невысокой численности стрептомицетов характеризуются присутствием в основном неокрашенных форм (Кузнецов, 1961; Андреюк и др., 1974; Зенова, Вустина, 1980). Дерпово-подзо- листые, торфяные, серые лесные почвы характеризуются относительно узким групповым спектром стрептомицетов (индекс группового разнообразия не превышает 0,33—1,16), доминирующими видами являются Slreptomyces albus, S. griseus, S. chromogenes (Павловича, 1978; Зенова, Вустина, 1979). В каштановой почве, сероземе и особенно черноземе групповой спектр значительно расширяется (индекс разнообразия 1,8) (Андреюк и др., 1974).
Селективное выделение актииомицетов, позволяющее подавить развитие в высевах на плотные среды грибов, бактерий и стрептомицетов, в последнее десятилетие дало возможность выделить и изучить ряд новых форм этих микроорганизмов (Преображенская и др., 1978; Buchell, 1982; Williams, Wellington, 1982).
Имеются сведения об определенной эколого-географпческой локализации представителей некоторых родов и видов актииомицетов. Так, культуры с мутовчатым ветвлением спороносцев встречаются в почве очень редко в количестве, не превышающем 0,27—1,37% от числа других актииомицетов. Обычно стрептовертициллы ограничены «рамками» 60° северной (и, вероятно, южной) широты (Конев, Цыганов, 1966; Конев PI др., 1974; Коиев, 1962). Представители гло- биспориновой группы стрептомицетов распространены повсеместно от Арктики до Антарктики (Szabo, 1974).
Сведения о распределении в почвах других родов актиномицетов в литературе ограничиваются единичными упоминаниями. Известна, например, приуроченность актиномадур к черноземам и каштановым почвам (Preobrazhenskaja et al., 1978). Представители рода Thermoactinomyces — космополиты (Cross, 1981а). Численность Thermoactinomyces vulgaris и Т. thalpophilus в почвах с нейтральным значением рН и хорошим водно-воздушным режимом достигает 107 спор/г субстрата. Споры термоактиномицетов вмываются в реки и ручьи, откладываются в илах озер и донных осадках морей (Cross, 1981в). Они чрезвычайно долговечны и сохраняют жизнеспособность сотни лет в сухой почве и свыше 2000 лет в осадках холодных анаэробных озер.
Подавляющее большинство работ по определению численности актиномицетов выполнено методом посева почвенных суспензий вглубь или на поверхность агаризованных питательных сред. Однако этот метод не дает информации о формах существования актиномп цетов в почве и их связи с субстратом Большинство колоний, очевидно, развивается из гпор, особенно если почва не обогащена питательным веществом (Mayfield et al., 1972). Поэтому для определения численности актиномицетов в почве применяют также непосредственное микроскопироваттпе почвенной взвеси (Аристовская. 1965) и окрашенной сугпеизии с измерением д^ины нитей мицелия и подсчетом спор (Павловича, 1978). Кроме того, используется люминесцентная микроскопия почвенной суспензии с учетом длины обрывков мицелия толщиной менее 1 мкм (Звягинцев н др., 1976а). Существуют попытки определения биомассы мицелия актиномицетов л почве.
Биомасса 1 м мицелия (принимая удельную плотность микроорганизмов равной 1 г/см3, диаметр гиф 0,5 им. и содеожание воды Б клетке 80%) составляет 3,9 10~8 г (Полянская, 1978) Отсюда биомасса мицелия актнномицетов равна 2,5• 10~5 г на 1 г почвы (640 м мицелия в 1 г почвы), т е ^актиномицетьт составляют 1.5% от биомассы всех почвенных микроорганизмов (Полянская, 1978)
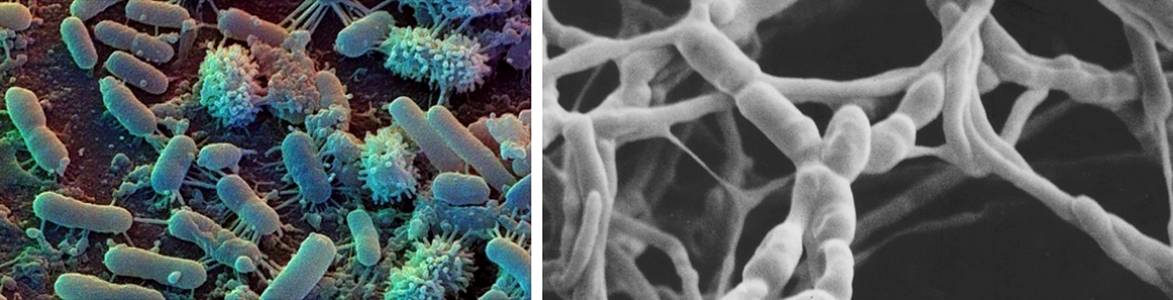
Первые светооптические наблюдения показали, что многие актн- номицеты, развиваясь в почве, могут образовывать воздушный мицелий и споры (Knbienna. Renn, 1935: Erikson, 1947; Pfenning, 1958) Однако способность к образованию мицелия может быть редуцирована (Красильников, 1958) С помощью педоскопов (Аристовская. 1965), прямой электронной микроскопии почвенной суспензии (Никитин и др., 1966), флюоресцентпой микроскопии (Звягинцев, 1959) сканирующей электронной микроскопии (Williams, Cross. 1971: Mayfield et al., 1972; Gray, Williams, 1971) в почве удается обнаружить спороношение актиномицетов, обычно наблюдаемое в лабораторных Условиях. Однако оно встречается реже, чем можно было бы ожидать, исходя из повсеместной распространенности этих организмов. Экспериментальные данные по дифференцированному учету спор и мицелия при посеве с прогреванием или встряхиванием почвенной суспензии (Крючков, 1973; Полянская, 1978; Михайлов, Зе- нова, 1980; Skinner, 1951; Mayfield et al., 1972), при регистрировании флюоресцирующего материала, накапливаемого в гифах (Williams et al., 1970), и цейтраферной съемке (Lloyd, 1969) свидетельствуют о доминировании спор в популяциях актиномицетов в почве. Доля колоний актиномицетов в высевах из почвы, образованных спорами, составила 50—60% от всех колоний (Mayfield et al., 1972).
Применение люминесцентной и сканирующей электронной микроскопии для исследования актиномицетов в почве
Применение люминесцентной и сканирующей электронной микроскопии для исследования актиномицетов в почве позволило выявить активно растущий мицелий, ограниченный микрозонами с высокой концентрацией органического вещества (Williams et al., 1970; Mayfield et al., 1972; Williams, Khan, 1974). Убедительным доказательством активной жизнедеятельности актиномицета в почве является образование гранул антибиотика гелиомицина в мицелии Streptomyces olivocinereus, выявленное с помощью люминесцентной микроскопии (Звягинцев и др., 19766).
Внесение в нестерильную почву суспензии клеток Streptomyces olivocinereus и S. lanatus и наблюдение за их развитием с помощью прямой микроскопии, а также дифференцированный учет вегетативных клеток и спор с помощью авто- и иммунофлюоресценции, позволили сделать вывод о стабилизации плотности внесенных популяций на определенном уровне в течение длительного времени. Подобная стабилизация свидетельствует о динамическом равновесии между стадиями активного роста и покоя в популяции (Полянская, 1978; Михайлов, Зенова, 1980; Звягинцев и др., 1981).
Актиномицеты — медленно растущие организмы (Flowers, Williams, 1977). Доминирование актиномицетов в субстратах связано обычно с наличием полимеров — крахмала (Гузев и др., 1981), хитина (Williams, Robinson, 1981), кератина (Noval, 1957), нефтепродуктов (Зенова, Красильников, 1968; Нестеренко и др., 1979). Продуцируя целый ряд гидролитических ферментов, актиномицеты осуществляют функции разложения таких устойчивых полимеров, как целлюлоза, хитин, ксилан, лигнин, гуматы.
Способность разлагать хитин — наиболее изученное свойство почвенных стрептомицетов. Из многих культур выделены внеклеточные хитиназы (Skujns et al., 1970; Тиунова и др., 1976а, б). Обогащение почвы хитином и клеточными стенками грибов ведет к увеличению относительного числа стрептомицетов (Mitchell, Alexander, 1962; Williams et al., 1970). В кислых подстилках и почвах, где продуктивность грибов составляет до 30 кг/га ежегодно, стрептомицеты играют главную роль в разложении грибного мицелия. От начальных до более поздних стадий разложения хитина в этих почвах повышается значение рН за счет выделений актиномицетами метаболитов, при этом отмечается сукцессионная смена ацидофильных актиномицетов нейтрофилами (Williams, Robinson, 1981).
Устойчивость актиномицетов в культурах и в почве к действию токсических агентов, таких, как фумиганты и инсектициды, может свидетельствовать об их использовании в качестве источников питания. Стрептомицеты весьма устойчивы к производственным дозам вносимых в почву гербицидов анилинового и фенольного типа, три- азинов, замещенных мочевины (Anderson, 1978). В культуре стрептомицеты способны разлагать устойчивые пестициды — ДДТ, триази- новые и бензокислотные гербициды (Anderson, 1978). Отдельные виды Nocardia окисляют некоторые феноксиалкалоидные кислоты (Taylor, Wain, 1962),виды Nocardia и Streptomyces — бипиридилы (Anderson, 1978).
( В литературе неоднократно отмечалось участие актиномицетов в разложении и синтезе гумусовых веществ в почве, т. е. их существенная роль в процессах почвообразования и создании почвенного плодородия (Plotho, 1951; Теппер, 1976; Орлов, 1974). Имеются сведения об использовании ими полифенолов гуминовых кислот в присутствии доступных источников углерода (Szegie, Gulyas, 1968). Отдельные представители рода Nocardia и Micromonospora способны окислять гуматы, принимая участие в минерализации гумусовых веществ в почве (Теппер, 1976).
Актиномицеты образуют темноокрашенные пигменты — меланины, являющиеся предшественниками гумусовых веществ в почве (Кононова, 1963; Орлов, 1974; Малама и др., 1975; Зенова, 1977; Лях,1981). Предполагается, что трансформация в гумусовые вещества происходит путем отщепления аминокислотных радикалов и измене» ния уровня окисленности пигментов.
Актиномицеты осуществляют влияние на азотный баланс почвы
Представители рода Frankia формируют ассоциации с небобовыми растениями (ольха, облепиха и др.). Растение в ответ на инвазию актиномицета образует клубеньки на корнях. В этих условиях акти- номицет начинает интенсивно фиксировать атмосферный азот (100— 300 кг N/ra в год). В настоящее время с несомненностью доказана возможность фиксации азота in vitro актиномицетами рода Frankia (Gauthier et al., 1981; Torrey, 1978).
Одна из характерных особенностей, благодаря которой актиномицеты стали предметом интенсивных исследований во всех странах мира,— их способность к образованию антибиотиков. Известно, что в лабораторных условиях актиномицеты способны синтезировать множество антибиотиков и других биологически активных веществ. В последнее время появились несомненные доказательства образования антибиотиков в почве: в нестерильной необогащенной почве обнаружен актиномицин, образуемый Streptomyces antibioticus (Stevenson, 1954), гелиомицин, образуемый Streptomyces olivocinereus (Звягинцев и др., 1976), антрахиноновые антибиотики, образуемые Streptomyces diastatochromogenes (Kaspari, 1973). Очевидно, в нестерильной почве существуют микрозоны, достаточно обогащенные питательным веществом, обеспечивающим активное развитие актиномицетов, синтезирование и функционирование антибиотиков.
На основе метода дифференцированного учета актиномицетов-ан- тагонистов, внесенных в нестерильную почву, показано (Михайлов, 1982), что антагонизм может проявляться непосредственно в почве при высоком уровне внесения (108—109л клеток на 1 г почвы) популяций. Плотность популяции актиномицета, подвергающейся действию популяции антагониста, резко падает с первых суток опыта, и уровень стабилизации ее оказывается на несколько порядков ниже уровня стабилизации других популяций. Однако это не приводит к гибели популяции актиномицета, подвергающейся действию антагониста. При совместном внесении пары популяций актиномицетов-ан- тагонистов в нестерильную почву на уровне 106 клеток на 1 г почвы плотность обеих популяций актиномицетов всегда ниже, чем при раздельном внесении (Михайлов, 1982).
Вопрос об экологической роли антибиотиков в почве широко обсуждается в литературе (Gottlieb, 1976; Калакуцкий, Агре, 1977; Demain, 1980).
Потенциал функциональных проявлений актиномицетов в природе далеко не исчерпан. Интереснейшие возможности актиномицетов открываются при исследовании их физиологических проявлений в ассоциациях с другими организмами (Калакуцкий, Агре, 1977; Зе- нова и др., 1980). Разработка методов исследования поведения актиномицетов в ассоциациях расширит наши познания в области функционирования актиномицетов в природе.
ЛИТЕРАТУРА
Андреюк Е. И., Владимирова Е. В., Коган С. В. Актиномицеты почв юга европейской части СССР и их биологическая активность. Киев: Наук, думка, 1974. 143 с. Аристовская Т. В. Микрофлора подзолистых почв. М.: Наука, 1965. 187 с. Гузев В. СВызов Б. А., Звягинцев Д. Г. Амилолитическое микробное сообщество в черноземе.— Микробиология, 1981, т. 50, вып. 4, с. 698—699. Егорова С. В. Микрофлора темнокаштановых почв.— В кн.: Микрофлора почв южной части СССР. М.: Наука, 1966, с. 3—24. Жданникова Е. Н. Микробиологическая характеристика торфяно-болотных почв Томской области.— В кн.: Заболоченные леса и болота Сибири. М.: Изд-во АН СССР, 1963, с. 170—182. Звягинцев Д. Г. Адсорбция микроорганизмов почвенными частицами: Автореф. дис. ... канд. биол. наук. М.: МГУ, 1959. 34 с. Звягинцев Д. Г., Кожевин П. А., Малахов В. В. Экологические проблемы в почвенной микробиологии.—Журн. общ. биологии, 1976а, т. 37, № 5, с. 691—706. Звягинцев Д. Г., Виноградова К. А., Ефременкова Л. М. Прямое микроскопическое выявление в почве актиномицета — продуцента люминесцирующего антибиотика.— Микробиология, 19766, т. 45, вып. 2, с. 337—341. Звягинцев Д. Г., Зенова Г. Л/., Михайлов В. В. Динамика популяции Streptomy- ces lanatus в сероземе и дерново-подзолистой почве в зависимости от формы и плотности внесения. —Биол. науки, 1981, № 6, с. 98—101. Зенова Г. М. Меланоидные пигменты актиномицетов.— Биол. науки, 1977, № 7, с. 4-13. Зенова Г. Л/., Вустина Т. П. Динамика численности актиномицетов в подстилках и дерново-подзолистой почве под березняком и ельником.— Вестн. МГУ. Сер. 17. Почвоведение, 1979, № 2, с. 55—58. Зенова Г. М., Вустина Т. П. Актиномицеты в песчано-пустынных почвах и сероземах Туркмении.— Вестн. МГУ. Сер. 17, Почвоведение, 1980, № 1, с. 39—43. Зенова Г. М., Красилъников Н. А. Актиномицеты, разлагающие углеводороды парафинового ряда.— Микробиология, 1968, т. 37, вып. 5, с. 870—875. Зенова Г. М., Лихачева А. А., Суетин О. СКалакуцкий Л. В. Совместная культура актиномицета и зеленой водоросли.— Биол. науки, 1980, № 5, с. 73—78. КалакуцкийЛ. ВАгре Я. С. Развитие актиномицетов. М.: Наука, 1977. 286 с. Клевенская И. Л. Микрофлора черноземов Сибири.—В кн.: Микрофлора почв северной и средней части СССР. М.: Наука, 1960, с. 250—273. Конев Ю. Е. Условия для выявления мутовчатого строения спороносцев у актиномицетов.— Микробиология, 1962, т. 31, вып. 2, с. 265—270. ^ Конев Ю. Е., Цыганов В. А. Мутовчатые актиномицеты-продуценты полиеновых антибиотиков.— Микробиология, 1966, т. 35, вып. 4, с. 612—622. - Конев Ю. Е., Барашкова Н. П., Шенин Ю. Д. Новые мутовчатые актиномицеты, образующие гексаеновые антибиотики.— Микробиология, 1974, т. 43, вып. 4, с. 659-665. Кононова М. М. Органическое вещество почвы. М.: Наука, 1963. 314 с. Красильников Н. А. Микроорганизмы почвы и высшие растения. М.: Изд-во АН СССР, 1958. 463 с. Красильников Н. АКореняко А. М., Артамонова О. И. Распространение акти- номицетов-антагонистов в почвах.— Микробиология, 1953, т. 22, вып. 1, с. 3-10. Крючков В. А. Метод определения состояния актиномицетов в почве.— Экология, 1973, № 1, с. 84—85. Кузнецов В. Д. Актиномицеты красноземов Зеленого Мыса и их антагонистические свойства.— Антибиотики, 1961, № 26, с. 781—786. Лях С. П. Микробный меланиногенез и его функции. М.: Наука, 1981. 274 с. Малама А. А., Храменко Г. В., Орлов Д. С., Юхнин А. А. Элементный состав и инфракрасные спектры меланиновых пигментов некоторых микроорганизмов.— Изв. АН СССР. Сер. биол., 1975, № 5, с. 766—768. Михайлов В. В. Взаимодействие популяций актиномицетов в почвах: Автореф. дис. ... канд. биол. наук. М.: МГУ, 1982, 21 с. Михайлов В. В., Зенова Г. М. Динамика и структура популяции Streptomyces lanatus в двух типах почв.— Микробиология, 1980, т. 49, вып. 6, с. 1011—1013. Мишустин Е> Н. Микроорганизмы и продуктивность земледелия. М.: Наука, 1972. 343 с. Мишустин Е. II. Ассоциации почвенных микроорганизмов. М.: Наука, 1975.107 с. Мишустин Е. Н. Развитие учения о методах почвенных микроорганизмов.— В кн.: Тез. докл. VI съезда ВОП. Тбилиси, 1981, т. 2, с. 161—162. Мишустин Е. Е., Мирзоева В. А. Соотношение основных групп микроорганизмов в почвах разных типов.—Почвоведение, 1953, № 6, с. 1—10. Никитин Д. ИВасильева Л. В., Лохмачева Р. А. Новые и редкие формы почвенных микроорганизмов. М.: Наука, 1966. 70 с. Новогрудский Д. М. Микробиологические процессы в почвах полупустынь,— Микробиология, 1946, т. 15, вып. 3, с. 177—186. Орлов Д. С. Гумусовые кислоты почв. М.: Изд-во МГУ, 1974. 333 с. Павловича Д. А. Актиномицеты Латвии. Рига: Зинатне, 1978. 194 с. Полянская Л. М. Популяция Streptomyces olivocinereus в почвах разных типов: Автореф. дис. ... канд. биол. наук. М.: МГУ, 1978, 22 с. Преображенская Т. П., Агре Н. СКалакуцкий Л. В. Новые формы актиномицетов.— Успехи микробиологии, 1978, вып. 13, с. 84—105. Рыбалкина А. В., Кононенко Е. В. Активная микрофлора почв.— В кн.: Микрофлора почв Европейской части СССР. М.: Изд-во АН СССР, 1957, с. 174—258. Нестеренко О. А., Касумова С. АКвасников Е. И. Ассимиляция углеводородов микроорганизмами рода Nocardia группы «rhodochrous».— Микробиол. журн., 1979, т. 41, вып. 1, с. 110—114. Теппер Е. 3. Микроорганизмы рода Nocardia и разложение гумуса. М.: Наука, 1976. 197 с. Тиунова Н. А., Пириева Д. А., Фениксова Р. В., Кузнецов В. Д. Образование хитиназы актиномицетами в погруженной культуре.— Микробиология, 1976а, т. 45, вып. 1, с. 246—248. Тиунова Н. А., Пириева Д. А., Фениксова Р. В. Образование и свойства хитиназы Act. kursanovii.— Микробиология, 19766, т. 45, вып. 4, с. 543—646. Чу лаков Ш. А. Влияние различных способов обработки почв на динамику микробных процессов.— Тр. ИНМИ АН СССР, 1960, вып. 7, с. 249—259. Anderson J. R. Pesticide effecls on non-target soil microorganisms.— In: Pesticide microbiology / Ed. J. R. Hill, S. J. L. Wright. L. etc.: Acad, press, 1978, p. 312-ч- 533. Buchell M. E. Microbiological aspects of the discovery of povel secondary meta* bolites.— Topics Enzyme Ferm. Biotechnol., 1982, vol. 3, p. 32—67. Cross T. Aquatic actinomycetes.—J. Appl. Bacteriol., 1981a, vol. 50, p. 397—423. Cross T. The monosporic actinomycetes.— In: Prokaryotes / Ed. M. P. Starr et al. B. etc.: Spring.-Verl., 1981b, vol. 2, p. 2028—2090. , Davis F. L.j Williams S. T. Studies on the ecology of actinomycetes in soil. 1. The occurence and distribution of actinomycetes in a pine forest soil.— Soil. Biol, and Biochem., 1970, vol. 2, p. 227—238. Demain A. L. Do antibiotics function in nature? — Search, 1980, vol. 11, N 5, p. 148-151. Erikson D. Studies on some lakemud strains of Micromonospore.— J. Bacteriol., 1947, vol. 41, p. 277—300. Flowers Т. H., Williams S. T. The influence pH on the growth and viability of neutrophilic and acidophilic streptomycetes.— Microbios. 1977, vol. 18, p. 223— 228. Gauthier D., Diem H. G., Dommergues Y. In vitro nitrogen fixation by two acti- nomycete strains isolated from Casuarina nodules.— Appl. Microbiol. Environ., 1981, vol. 40, N 1, p. 306-308. Gottlieb D. The production and role of antibiotics in soil.— J. Antibiot., 1976, vol. 29, N 10, p. 987—1000. Gray T. R. G., Williams S. T. Microbial productivity in soil.— In: Microbes and biological productivity / Ed. D. E. Hughes, A. H. Rose. Cambridge: Univ. press, 1971, p. 225—286. Kaspari H. Untersuchungen iiber Bildung und Aktivitat von Streptomycetenanti- biotika im Boden. I. Bildung von anthrachinon-antibiotika im Boden.— Zentr.-Bl. Bakteriol. Parasitenk., Infektionskrankh. und Hyg. II. Abt. Orig. B, 1973, Bd. 128, S. 764—771. Khan M. R., Williams S. T. Studies on the ecology of actinomycetes in soil. VIII. Distribution and characteristics of acidophilic actinomycetes.— Soil Biol, and Biochem., 1975, vol. 7, p. 345—348. Kubienna W.f Renn С. E. Micropedological studies of the influence of different organic compounds upon the microflora of the soil.— Zentr.-Pl. Bakteriol. Parasitenk. Infektionskrankh. und Hyg. II. Abt. Orig. B, 1935, Bd. 91, S. 267—292. Kuster K. Ecology and predominance of soil streptomycetes.— In: Actinomycetes — the boundary microorganisms / Ed. T. Arai. Tokyo etc., 1976, p. 109—121. Kutzner N. J. The family Streptomycetaceae.— In: Prokaryotes / Ed. M. P. Starr et al. B. etc.: Spring.-Verl., 1981, vol. 2, p. 2028—2090. Lacey J. Actinomycetes in soils, composts and fodders.— In: Actinomycetales: Characteristics and practical importance / Ed. G. Sykes, F. A. Skinner. L.; N. Y.: Acad, press, 1973, p. 231—251. Lechevalier II. A. Distribution et role des actinomycetes dans les eaux.— Bull. Inst. Pasteur., 1974, vol. 72, p. 159—175. Lechevalier H. A., Lechevalier M. P. Introduction to the order actinomycetales.— In: Prokaryotes / Ed. M. P. Starr et al. B. etc.: Spring.-Verl., 1981, vol. 2, p. 1915—1922. Lloyd A. B. Behavior of streptomycetes in soil.— J. Gen. Microbiol., 1969, vol. 56, p. 165—170. Mayfield C. /., Williams S. TRuddick S. /., Hatfield H. L. Studies on the ecology of actinomycetes in soil. IV. Observations on the form and growth of streptomycetes in soil.— Soil Biol, and Biochem., 1972, vol. 4, p. 79—91. Mitchell R., Alexander M. Microbiological processes associated with the use of chi- tin for biological control.— Proc. Soil Sci. Soc. Amer., 1962, vol. 26, p. 556—558. Noval J. The decomposition of keratin by microorganisms: Ph. D. Thesis. New Brunswick, 1957. 31 p. Okami Y. Antibiotics produced by actinomycetes.— In: Handbook of microbiology/ Ed. A. J. Laskin, H. A. Lechevalier. Cleveland: CRC press, 1973, vol. Ill, p. 717— 972. Pfenning N. Beobachtungen des Wachstumsverhaltens von Streptomyceten auf Rossi-Cholodney-Aufwuchsplatten 1m Boden.— Arch. Mikrobiol., 1958, Bd. 3i, S. 206—216. Plotho О. V. Weitere Untersuchungen zur Huminbildung der Microorganismen.- Ztschr. Pflanz. Dung., Bodenk., 1951, Bd. 55, S. 151. Preobrazhenskaja T. P., Sveshnikoua M. A., Terekhoua L. P., Chormonova N. T. Selective isolation of soil actinomycetes.— In: Nocardia and StreptomyQes / Ed. M. Mordarski, W. Kurylowicz, J. Jeljaszewicz. Stuttgart: G. Fischer Verl., 1978, p. 119—123. Skinner F. A. A method for distinquisching between viable spores and mycelial fragments of actinomycetes in soils.— J. Gen. Microbiol., 1951, vol. 5, p. 159—166. Skujns /., Pukite A., McLaren A. D. Chitinase of Streptomyces sp.: purification and properties.— Enzymologia, 1970, vol. 39, p. 353—370. Stevenson /. L. Antibiotic production by actinomycetes in soil demonstrated by morphological changes induced in Helminthosporium sotivum.— Nature, 1954, vol. 174, N 4430, p. 558—599. Szabo I. M. Microbial communities in a forest-rendzine ecosystem. Budapest: Acad. Kiado, 1974. 415 p. Szegie /., Gulyas F. Data on the humus-decomposing activity of some Streptomyces and microscopic fungi.— Agrohem. Talayt, 1968, vol. 17, p. 107—119. Taylor H. F., Wain R. L. Side-chain degradation of certain w-phenoxyal kanecar- boxylic acids by Nocardia coeliaceae and other microorganisms isolated from soil.— Proc. Roy. Soc. London B, 1962, vol. 156, p. 172—186. Torrey G. H. Nitrogen fixation by actinomycete nodulated angiosperms.— Bioscience, 1978, vol. 28, p. 586—592. Waksman S. A. The actinomycetes. Vol. I. Nature, occurence and activities. Baltimore: Williams, Wilkins Co, 1959. 327 p. Waksman S. A., Lechevalier H. A. The actinomycetes. Vol. III. Antibiotics of actinomycetes. Baltimore: Williams and Wilkins, 1962. 346 p. Williams S. T. Streptomycetes in the soil ecosystem.— Zentr.-Bl. Bakteriol., Para- sitenk. Infektionskrankh. und Hyg. I. Abt., 1978, Bd. 6, Suppl., S. 137—144. Williams S. Г., Cross T. Actinomycetes.— In: Methods in microbiology / Ed. J. R. Norris, D. W. Robbens. L.; N. Y.: Acad, press, 1971, vol. 4, p. 295—334. Williams S. Т., Khan M. R. Antibiotics — a soil microbiologists viewpoint.— Post, hig. i med. dosw., 1974, wol. 28, s. 395—408. Williams S. Т., Robinson C. S. The role of streptomycetes in decomposition of chi- tin in acid soils.— J. Gen. Microbiol., 1981, vol. 127, p. 55—63. Williams S. Т., Wellington E. M. H. Principles and problems of selective isolation of microbes.— In: Bioactive microbial products, search and discovery / Ed. J. D. Bu'Lock et al. L. etc., 1982, p. 9—26. Williams S. THatfield M. L., Mayfield C. Y. New methods for the observation of actinomyceten growing on artificial and natural substrates.— In: The actinomycetes / Ed. M. Prauser. Jena: Gustav Fischer Verlag, 1970, p. 379—399. |











Актиномицеты

К содержанию книги: Почвенные организмы как компоненты биогеоценоза